Now, before continuing with the epochal survival benefits inherent in this new functionality, the bottom-up engineer must be reminded that at this point, no matter how cleverly this vertebrate eye arrangement can resolve coherent visual fields from their backgrounds, the emotive complex in our vertebrate neural array is still being presented with photonic sensation that has no emotive quality to it. And even if this marks the graduation of organic vision from the signaling of shadows to the epochal imaging of light itself, the emotive complex in our organism still does not have the cues necessary to steer away or steer toward these emergent visual fields.
So, as has been the case ever since the imagination scenario began with our early primordial metazoan, it is the emotive complex, and its guiding feedback of perceptual diffraction, that will be the shaping force for the early vertebrate optic tectum and the imaging process evolving in our Devonian organism.
If the contrast extraction/boundary resolution process represented our organisms’ graduation from shadows to the imaging of light, the determination of those cues needed to differentiate between the attractive and aversive characteristic of individual visual fields would represent the post-graduate advancement as the neural array progresses from sensation to perception. The sensation of contrast states in the photonic signaling provided by the vertebrate eye is only the beginning of the story. The perception of the visual world will require a number of neuroanatomical adaptations as well.
It was the development of the neural map structure that allowed the transition from the compound eye to the dual-lateral vertebrate eye, and unlike the compound eye arrangement, which required only one neural map for its production of global associations, the vertebrate eye system will require two neural maps at first, one for each lateral eye. The photonic sensory complex of Cambrian metazoans is now evolving into the optic tectum of post-Cambrian vertebrates, and the requirement for dual neural maps supporting each eye will promote a hemispheric separation of the optic tectum into two co-existent lobes.
With this hemispheric separation of what has been characterized as the photonic sensory complex, comes an equally important anatomical adaptation. Although it would seem that developing an ability to first objectify those coherent image fields that have emerged apart from their background by the optokinetic process would be the next important step in the imaging process, it is the discrimination between aversive and attractive qualities of those emerged image fields that remains a primary imperative in that imaging process. Objectification implies cognition, but our early vertebrate neural array is not there yet. In between the pure sensation of Cambrian metazoans and the cognitive behavior of the much more sophisticated tetrapods, lies the neural process of emergent perception.
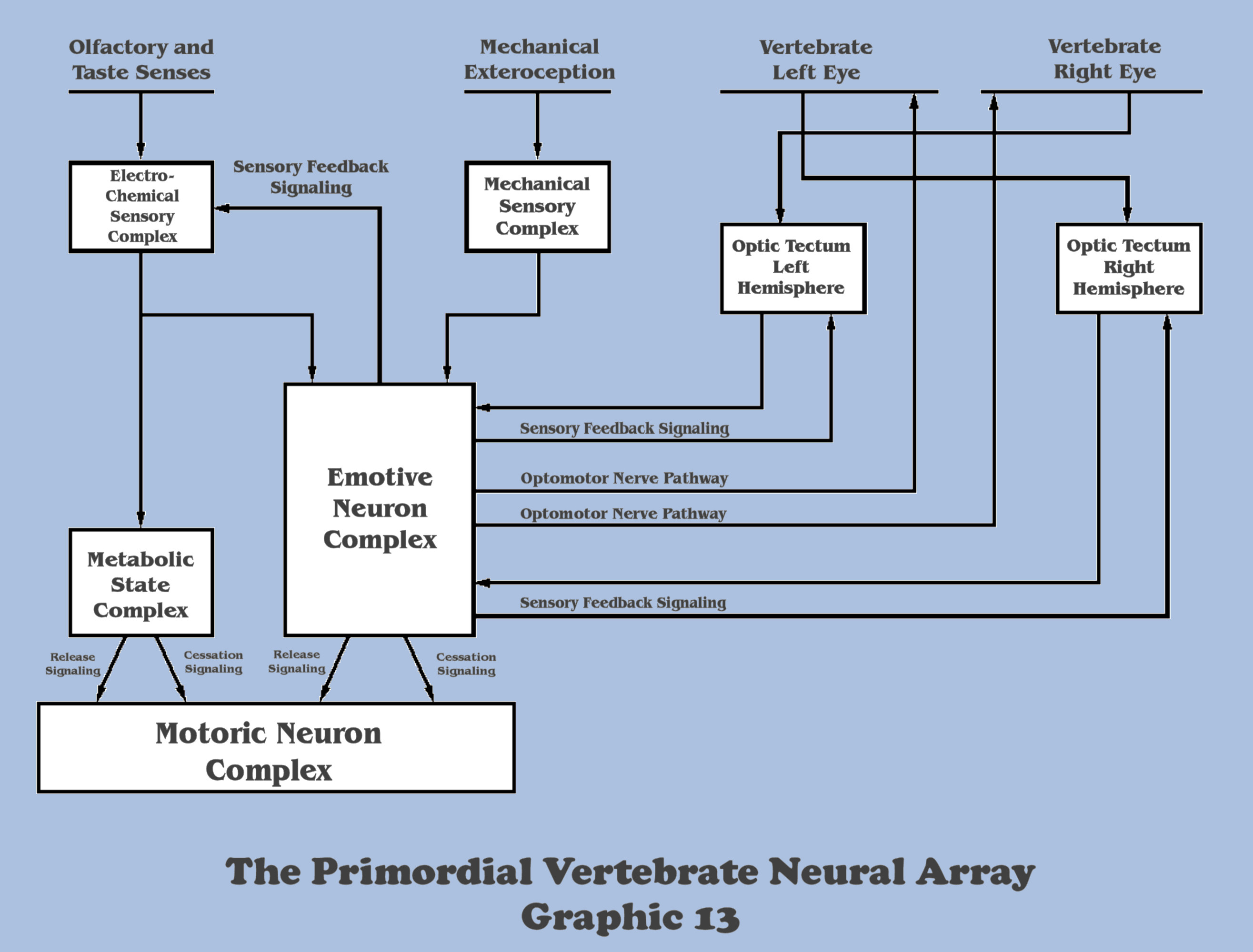
With the incipient evolution of the neuroanatomical processes to create the production of segregated image fields by the optokinetic response, further anatomical adaptations that begin the progression to emergent perception shall center on the emotive complex of the organisms’ neural array once more. Graphic 13 illustrates the specializations of the dual optic hemispheres possessing individual neural maps in the changing neural anatomy, and something more.
Because the sensory imperative to abstract the emotive character of any imagery comes before the much more complex processes of objectification, a feedback channel must be established between the imaging processes of the optic tectum and the emotive complex. And because of the hemispheric segregation of the tectum, there must be a separate but interdependent feedback channel for each hemisphere.
Also in Graphic 13, we see additional new components. With the wide angle coverage of compound eyes being replaced with a narrower field view in vertebrate eyes, there will now be a pre-eminent channel required in all vertebrates to actively control the gaze angle of vertebrate eyes. Graphic 13 illustrates the conceptualization of the optomotor nerve pathways controlling the muscles of the eyes themselves.
Returning to the optokinetic response process in the free swimming vertebrate, our imagination scenario seems to be at the identical dilemma that had developed when the sensory complex of the primordial metazoan produced a dangling direction vector having no emotive value for the emotive complex to respond to. In our current imagination scenario, once the resolved image fields have emerged via their motion against the background in the optokinetic process, the vertebrates’ neural array will not have the luxury to develop any objectification of their characteristics because each hemisphere will be in a literal race for survival with the other hemisphere. During the normal course of swimming, the emotive complex must know, before any objectification occurs, in which hemisphere an aversive predator might emerge in time for the organism to set up the next thrust cycle in its locomotion.
Recall that the optokinetic process has been synchronized to the cyclic phases of an organisms’ fin strokes, and the entire procedure began with the propulsive phase of the thrust stroke. It was then during the subsequent refractory phase of the fin stroke that all of the imaging of the signals from the eyes must occur, in order for the emotive complex to immediately trigger an avoidance maneuver away from a predator if need be on the next propulsive stroke of the fins. For this, the emotive complex must know in which hemisphere the predator image has emerged, so as to signal the proper direction to take in its evasive maneuver away from danger.
There is no reason not to believe that each hemisphere would follow identical processes in the determination of this first situational imperative. As stated in the previous few paragraphs, the functional nature of this imperative determination is presently apparent, but what is not so obvious is the neurological basis that Nature would craft to implement this determination.
The evolution of the optokinetic apparatus has resulted in a mechanism by which the image fields of moving objects in the visual fields of swimming vertebrates “pop-out” from their background, and it is just these emergent fields that are presented to the image processing array of the organisms’ optic tectum, separate from that background. The determination of the emotive character in these emergent fields could be implemented by a phylogenetically established decision filter, a filter whose decision criterion is based on any number of possible discernable aspects of the image fields themselves, such as, in its most simplest case, the evolution of an ability to discriminate the angle that an emergent “pop-out” field subtends relative to the entire visual field. The decision criterion might set a certain relative angle value at which the subtended determination would trigger an avoidance reaction in the organism.
Behaviorally, this emotive response mechanism seems straightforward, but the neurological processes required to bring this behavior about are not so simple. Nature is still inexperienced with the new task of imaging light, and such a simple emotive adaptation would reveal other selective pressures. In addition, this simple response mechanism results in an avoidance reaction to any visual field triggering the decision criterion, and does not allow for an emotive response to attractive mates or prey.
The determination of emotive responses to aversive or attractive characteristics requires the spatial differentiation of emergent image fields in the neural map, which now calls for a more sophisticated image analysis capability than that of the simple Cambrian boundary resolution process. Survival of the free swimming vertebrate will require a new ability, that of cue derivation from these emergent image fields, an operation somewhat like the local contrast extraction to boundary resolution affair that resulted in the neural structures to begin imaging in the first place, but the distillation of cues will require a new imaging mechanism in the neural maps of vertebrates, an abstraction process which will bring about map wide state synthesis. This new selection pressure will promote adaptations to the metabotropic state synthesis enterprise, nudging it from the production of simple ionotropic persistences toward complexities approaching the expression of true memory.
The dialog has illustrated how neural assemblies have utilized metabotropic processes to abstract change in the instantaneous signaling of exteroceptive inputs, or even change in the instantaneous signaling within the neural array itself. And Nature would no doubt continue with this technique in the evolving stages of vertebrate image analysis following the production of emergent image fields during the optokinetic process. But the previously illustrated examples of metabotropic abstraction in past discussions involved the biasing of instantaneous signaling at a single dendrite, or, with second order state synthesis, within the confines of a local neural assembly. With the current adaptations of emerging visual fields produced by the optokinetic response, the organism now requires an elementary abstraction facility across a global population of non-interconnected neural coalitions.
Recall that in the resolution of the dangling direction vector, the mechanism signaling change was effected by the simultaneous comparison of two temporally disjoint local state abstractions, local abstractions by virtue of being formed by first and second order metabotropic persistences in the simple phasic ensemble signaling the composite direction vector itself. But with the requirement to implement cue derivation from a global population of map-wide neural signaling, what is needed now is not the abstraction of change across the temporally disjoint states of a local phasic assembly, but the differentiation of cue domains across the spatially disjoint states of a global neural map.
The imaging of vertebrate photonic signaling fundamentally relies upon the ability of the neural array to maintain topologic coherence of the visual field across sequential imaging steps. The first step of that imaging, the “pop-out” of emerged image fields in the neural maps of the optic tectum produced by the optokinetic response machinery, preserves that topologic coherence. Now, for the bottom-up engineer, if neural maps can be conceived of as the functional part of a “procedural language” for global definitions in an “imaging semantic”, what Nature needs following that conceptualization are syntactic forms in which to maintain this topologic coherence across subsequent steps of that imaging semantic.
Given Natures’ inexperience with the imaging of light at this point in the evolution of our primordial vertebrate, neural adaptations which express these syntactic transformations would probably have to come incrementally, but certainly, with the demands of survival pressuring those adaptations, the first syntactic form in imaging analysis that Nature would have to learn would be those transformations that yield spatial invariance.
When Joseph Fourier developed his equations for an analytic theory of heat transfer in 1822, he established the semantics for domain transformations between a time series and a frequency series. Since much of life’s dynamics can be expressed in the language of periodic series, Nature would rediscover this meme of domain transformations many times in the multitude of adaptations contributing to the evolution of organic nervous systems.
And although these fundamentally powerful transformations between time and frequency series would not find their way into Natures’ designs until the development of more complex cerebral structures, there was a specific aspect of Fourier’s invention which Nature did adopt for the initial image analysis in our primordial vertebrate.
A fundamental concept expressed in the Fourier series is that of functions, which are localized in the frequency domain, which have definitions that are functionally spread out (globalized) in the time domain.
Using this conceptualization, perhaps Nature could develop those syntactic expressions which inversely describe map-wide characteristics of the global neural map and transform them into local discriminations, or cues, which could be subjected to next-stage differentiation.
But unfortunately, Nature has not as yet determined what local cues in the global image have survival value for this discrimination process.
So, before Nature could phylogenetically define those local cues having higher survival value, selective pressures would lead to the development of domain transformations independent of these local cues. And the first of those independent transformations would be the functional expression of spatial invariance between the sequential process steps in the imaging process.
Spatial invariance can initially be visualized by the bottom-up engineer in the context of our primordial vertebrate image analysis using a brief example. Recalling the optokinetic response process, where the perception of imagery began when the contrast patterns of a topologically cohesive field bounding the extents of an object, say, for example, a hungry predator, “pops-out” due to its relative motion against the optokinetically stabilized background, this still presented Nature with a significant task in image analysis.
This bounded field emerges from the retinal ganglionic structure onto the neural map of a particular hemisphere of the optic tectum at a spatial location which could be anywhere relative to the global center of the retinotopic map.
In the example analysis, the neural array will require an ability to shift the local center of the emerged field onto the global center of the (perhaps secondary cue analysis) neural map, which is a transformation from the retinotopic frame of reference to a frame that is spatially invariant relative to any particular emerged visual field. If the organism was to accomplish this translation mechanically, by shifting the gaze of its eye to center on the emerged field, the temporal contrast patterns of the retinal map would be lost, and the whole image analysis process would have to begin anew, wasting precious milliseconds by the hundreds (and the bottom-up engineer should also note that the feedback intensive and time-consuming process of focusing the optic lens in each eye for distance accommodation has not even entered into the image analysis endeavor as yet.)
The selective pressures of survival will certainly not allow Nature to waste this precious time in the neurological race to detect predators. The first capture of the instantaneous signaling of “pop-out” visual fields from the optokinetic process must be taken advantage of, with no time to waste. The behavioral product of this evolutionary imperative can be seen in extant pelagic organisms to this day. Perhaps there are those bottom-up engineers who have felt that somewhat creepy feeling one gets while observing fish (and especially sharks!) whose eyes rarely center on objects they might be focusing on.
Invariance – the property of remaining unchanged regardless of changes in the conditions of measurement – will certainly be invoked in a multitude of various spatial as well as temporal transformations well before Nature learns what specific retinotopically local cue derivations in emergent image fields have the most survival value. Although an image analysis scheme initially based on global transformations does not follow the classic (but questionable) approach that many neurologists use in modeling organic vision, (where the perceived visual space is deconstructed into local elemental features, such as bounded edges, which are subsequently built back up by a somewhat mysterious, hierarchical association mechanism into “objects” having presupposed cognitive sensibility), proceeding from a vision process capable of global transformations independent of specific constraints in the environment would represent Natures’ best strategy for phylogenetically resolving those retinotopically local visual cues with the most survival value.
And although Nature would discover a number of various expressions for neurologic general invariance in the evolution of central nervous systems, the dialog will initially focus on this first expression of spatial invariance to illustrate the subsequent path leading to the phylogenetic resolution of those local cues in the neural analysis of imagery that Nature will devise, which ultimately provide the most survival value to our evolving primordial vertebrate.
It has always been the intention of this dialog to help prevent the bottom-up engineer from wading into the intellectual quicksand of hierarchical thinking, but the presentation of foundational concepts before the establishment of their rational significance is a difficult instructional method, placing the burden of comprehension mostly on the student. But when this burden is successfully carried out by the astute learner, it turns into the joy of scientific discovery, which should be the true ideological desire on the part of every instructor for those instructees. And since the Introduction, the Organon Sutra has followed this Aristotelian form of presentation before rational resolution in the dialog, which has placed a huge burden on the intellectual fluency of the bottom-up engineer so far, but even this will seem trivial as the discussions move from demonstrating organic sensation to neural perception, thence to behavioral cognition and ultimately to emergent intelligence, an ever increasing burden requiring the supreme exercise of scientific consideration on the part of the reader.
So the presentation of the foundational aspects supporting the discussion of a neurological process for primordial imagery and the movement from organic sensation to neural perception will begin the even more demanding requirement for intellectual clarity on the part of the bottom-up engineer, because the epochal evolution of the vertebrate eye is actually the culmination of many singular adaptations, each comprehensive on its own, which has caused the discussion to unfurl into an echelon of foundational concepts, an intellective layer cake with no intervening resolution to any rational significance between layers. At this point the bottom-up engineer can no longer be a spectator to the spinning plates act that has been the Organon Sutra to this point, she must come up on stage to keep the spinning plates themselves from crashing to the ground.
The title of this discussion – “The progression from Sensation to Perception” – is but one layer of that subjective layer cake, and formed below that was the conversation regarding the primordial imagery processes, below which the dialog introduced the functional requirement for cue extraction in that imagery, and the necessities of primal emotive determination required another, even deeper layer of foundational concepts which might lead to a neurological implementation of transformations with global invariance. And the frosting over this hyper-tiered cake is the conceptual presentation of the Second Fundamental Precept, the elementary abstraction of state in the organism.
Finally, at the risk of comically over-extending this layer cake analogy, the bottom-up engineer should not be surprised to find that the frosting separating the tiered layers of the cake will be formed from the very metabotropic processes that have been a subject of the Second Fundamental Precept all along.
So it is only fair that the dialog should minimize that increasing burden on the part of the bottom-up engineer, but this should not be done by relaxing the boot-camp, drill instructor discipline achieved so far, and certainly should not be done at the prospect of minimizing the joy of discovery that insightful students might find. What we have in this intellectually amorphous layer cake is the philosophical springboard into the even deeper conceptualizations which the dialog has been calling the Third Fundamental Precept. The layer cake that will be developed in that examination promises to be even more multi-tiered, with each layer marbleized and even jelly-filled. But take heart, as the bottom-up engineer will find universal resolution to all of the loose ends in the Organon Sutra tapestry following that last fundamental precept, as the final parts of the dialog close the circle on a unified conceptualization of emergent intelligence.
Although, alas, it might seem that the path from here to there stretches on interminally, let us take a conceptual knife and cut through the layer cake we have at hand, to get a different conceptual perspective on all of the interrelated mechanics that we have just described supporting the evolution of the early vertebrate eye. And let us unwind a weighty portion of that cargo of disconnected concepts that the bottom-up engineer has been carrying throughout the dialog ever since Discussion 3, where the question was raised: In what form would a systematic take to model the enigmatic “right-brain” thinking?
Let us use this imaginary knife to see how this concept of spatial invariance in particular, and conceptual invariance in general, will provide the first nebulous answer to that question, and how a concept of “right-brain” global invariance will complement an emerging “left-brain” predilection for localized objectification, as we begin to envision the engineered components of our artificial agent itself. The emergent behavior of artificial intelligence that we seek will be found in the neurological marriage between this globalized invariance that Nature is developing, and the localized objectification that Nature will be moving toward.
GO TO NEXT DISCUSSION
GO TO TOP OF DISCUSSION
Copyright © 2019 All rights reserved