When the early astronomers attempted a systematic description of all that they observed in the night sky, they encountered a baffling conundrum.
The ancient astronomers, having already cataloged the visible objects beyond the Earth, had embedded the non-moving stars in an imagined crystal sphere, whose center was placed at the stationary Earth. In addition to the celestial Sun and Moon, these ancient astronomers were also aware of stars that did not remain stationary. For these “wandering stars”, which we now call planets, additional transparent, crystalline spheres were envisioned, to which a distinct wanderer was affixed, with every sphere arranged in concentricity with each other. Each sphere had a different diameter in correspondence with the estimated distance of its wanderer from the Earth, and they rotated in a manner to match the apparent motion of their attached planets. And this whole system placed the Earth at the center of the Universe.
As the accuracy of their observations improved over that of the ancient astronomers, the early astronomers observed a baffling behavior in the real motion of the planets. A planets’ rate of progress across the starry heavens varied significantly with time, and in addition to that, this progress would periodically slow down, come to a halt, and then, inexplicably, the planet would move in a retrograde direction for a brief period from its normal motion.
Although there were numerous other celestial anomalies which added to these confounding observations, and contributed to an impetus to question the veracity of their model of “celestial spheres” and Earths’ centricity, there was a strong need to keep the Earth at the center of the Universe. Time and teachings had firmly established a dogma, reinforced by mysticism, mythology and superstition, which elevated the centricity of the Earth in the Heavens to a fact beyond criticism. Even Aristotle, one of the few philosophers with near unquestionable views for his time, discussed the idea that the Sun, rather than the Earth, was at the center of the Solar System, but dismissed the idea, following the very scientific observation that the stars do not reflect any relative change in position, as would be expected if the Earth did indeed move around the Sun. Unfortunately, his science was based on an understanding of astronomical distances that was much more limited than our modern understanding.
But this established dogma was at odds with the baffling observations of the planets. Now, around 150 B.C., the Greek Hipparchus developed a model to explain the irregular planetary motion while retaining the revered, pure circularity of movement demanded by the mystical ideas of perfect motion in heavenly bodies and the centricity of Earth in the Universe. He proposed orbits within orbits, which were called deferents and epicycles, which brought the observed data back into alignment with “scientific understanding”, and the perfection of heavenly motion was preserved once again.
This system of deferents and epicycles stood for more than a millennium with only a modification to the scheme by Ptolemy in the second century A.D. It took the inventions of Copernicus and Kepler in the fifteenth century to reshape the model into the more modern and astronomically correct heliocentric system, with the Sun at the center of the Solar System.
However, this modernization did not come about by any radically new data. The observational data of the early astronomers, though not as accurate as their 15th century counterparts, was not wrong. The more refined instruments of the Renaissance era still showed the planets performing their periodic retrograde dance, and yet the models of Copernicus and Kepler demonstrated no such thing.
The reason for the disparity of their observations with their conceptualization of the system as a whole could be characterized as apparent nonlinearity. Apparent nonlinearity is an observational artifact that occurs when sampling a multi-dimensional event space with a sub-dimensional apparatus. For the Renaissance astronomers, it was not until the dimensionality of the problem was increased, not until the observing telescope was placed on a moving Earth, that the observations of the orbiting planets made sense.
The confounded astronomers were trying to explain a system which had multiple dimensions of motion with observations made from an instrument that canceled out one of the dimensions of movement.
Apparent nonlinearity is certainly distinct from its abstract counterpart: mathematical nonlinearity. If we were to observe a stone falling, for example, and after observing the stone hitting the ground, we could mathematically characterize the stones trajectory before and after impact with the ground as a nonlinear event. And in this case we would indeed have nonlinear observations describing a truly nonlinear event.
But long before evolution would be able to deal with mathematical nonlinearity, Nature would have to build neural assemblies to reconcile apparent nonlinearity.
Although distinct from mathematical nonlinearity, apparent nonlinearity is no less a real phenomenon, as the early astronomers encountered as they tried to reconcile verified nonlinear observations with what their dogma demanded must be perfectly linear behavior. Indeed, even today, much of our scientific exploration of the multi-dimensional universe is carried out with observations conducted with sub-dimensional instruments.
And certainly Nature must have run into this very real phenomenon as she was developing the early nervous systems for the organisms possessing the uni-dimensional, linear mechanics of phasic and tonic neural assemblies.
This dialog concluded the previous discussion with Nature busily experimenting away with these two phylogenetically engineered neural aggregates within the evolving neural array, but as Nature was developing the complexes that were exposed to the environment, complexes required to steer the co-evolving locomotive axis, that experimentation must have had to contend with something new, for any organisms possessing these neural processes must have experienced an environment which presented all manner of multi-dimensional change to the uni-dimensional phasic and tonic assemblies that Nature first began experimenting with.
As with the early astronomers, the multi-dimensional stimuli of the environment must have resulted in apparent nonlinear input to these linear, uni-dimensional neural processes, causing formerly functional translations of phasic assemblies to become indeterminate, and tonic feedback circuits to wobble and drive out of resonance as previously linear reactivations fizzled.
The inexhaustible variety of the environment means that it will present situations that have never been encountered before, either in the phylogeny of a species or in the life of an individual organism itself. How does an organism come to have mechanisms that can form a relevant response to a previously never experienced situation? At the level of the neural array, these mechanisms can only be developed phylogenetically, but as bottom-up engineers, we can be more explicit, after having learned the lessons that Nature acquired only through time-consuming experimentation. And ultimately, we must have an understanding of these early lessons in order to comprehend the seismic evolutionary changes that will occur when whole clades of evolving species encounter environments which change within generations, an environmental instability for which the genome can no longer orchestrate phylogenetic adaptation. As bottom-up engineers, we must be ready to re-direct our conceptual paradigms to reflect this inevitable tectonic shift in the design of our artificial agent.
So it is not hard to envision that this apparent nonlinear stimuli would create a selection pressure for the next inventive neural arrangement that Nature was to take advantage of, and where the bottom-up engineer will find our next lesson. But we left off the last discussion with just a general notion of the first lesson that Nature had to reconcile for its solution to intentional motility. In the imagination scenario for our organism possessing the single locomotive axis, Nature discovered that motile strategies could not follow a singular “generic” form or sequence, beginning with the induction of a locomotive scheme triggered by a metabolic “need”, followed by various motile executions guided by sensory signals from certain electro-chemical exteroceptors to satisfy that need, and finally extinguished by some metabolic signal of satisfaction. Of course, Nature would discover and take advantage of “opportunity strategies”- those motility strategies that are trigged by attractive signals originating from the environment, but the induction and extinction of the motility strategy itself still originates with the neural complexes that signal metabolic state.
Because the environment also presented dangers as well as attractions, all motility strategies were eventually polarized by the emotive complex which evolved in response to the reality that aversive mobility strategies must be triggered by signals originating in the environment, and not by the organisms metabolic state, and just as importantly, must be extinguished by signals that originate in the environment as well. And in between these two signaling events there must be a mobility strategy which could vary in accordance with the variety of aversive conditions.
This lesson most certainly was learned by Nature as far back as the primordial era the Organon Sutra has characterized as the early orientation stage of neural array adaptation. And it is hoped that the bottom-up engineer is also coming to understand that the same selective pressures which evolved the emotive complex would also drive the development of a feedback mechanism between the emotive complex and the sensory complex, in addition to developing those specializations in the sensory complex that would allow for survival enhancing efficiencies in the sensation of the environmental cues that guide the execution and extinction of mitigation strategies. This feedback mechanism would develop a functionality the Organon Sutra has termed “perceptual diffraction”, which is the biasing of the sensory complex to “see” the environment differently, depending upon the aversive/attractive feedback signaling of the emotive complex. However, the dialog had to suspend the discourse on this feedback mechanism, because as with all resonant assemblies, the feedback structures could not fully develop until Nature had fashioned the exteroceptive specializations that would be modulated by them, and that story revolves around Natures reconciliation of apparent nonlinearity.
And before proceeding, the dialog wants to point out that with the lesson of intentional motility, it is ultimately important for the bottom-up engineer to understand the intentional nature of the motoric complex in which the selective pressures are shaping, as the emotive complex develops in our evolving neural array. This intentional nature can best be spelled out by emphasizing to the bottom-up designer that in every motoric complex, the neural commands to cease an attractive, and more importantly, an aversive motoric sequence must be just as intentional as the command to initiate the sequence. Nature had to be just as specific in evolving the mechanisms to cease motoric strategies as she was in fashioning the mechanisms that initiate them.
The reason is because the sensory signals that initiate a motoric strategy may be different from the sensory signals that extinguish it.
The consequence for this (thinking on the part of the bottom-up engineer, and expression on the part of evolution), is to begin to bring the design of motoric activity above a mere consequential chain of steps, to elevate the design to a level above mere reflex action. Although intentional motility implies a “cognitive” aspect which our primordial neural array certainly does not exhibit now, it provides a mechanism or interface for the future “cognitive complex”, a complex that Nature will necessarily evolve, as we shall see.
There are additional consequences to this. Intentional behavior in the motoric complex will pressure the formation of foundational circuitry that will lead to the neural processes engendering elementary abstraction in the sensory complex. Although this may not be readily apparent at this stage in neural organization, perhaps the bottom-up engineer can grasp how the intentional nature of locomotion will ultimately lead to neural complexities that process if/then situations, if only on a most basic spatial level, with no temporal aspect at this point.
From an anatomical perspective, our primordial neural array at this point in the early evolution of the orientation stage might comprise a quantity of neurons on the order of tens or hundreds of thousands, depending on the degree of neural maintenance required for the organisms’ metabolic processes and the complexity of the muscular system empowering the locomotive axis. Yet even with this modest neurological size, the array at this stage is already exhibiting the primal behaviors of intentional motility which will lead to the more sophisticated “cognitive” functions, and developing sensory complexes with the progenitors of elementary abstraction. It is no wonder that hierarchical design is stillborn at even this stage in its attempt to produce anything like this level of modest complexity.
Although there is no clean break between Natures’ lesson on the reconciliation of intentional motility, and the subsequent lesson on the reconciliation of nonlinearity, we will leave the unresolved development of the feedback between the emotive complex and the sensory complex to a later discussion as the dialog directs the bottom-up engineers’ attention to Natures’ development of the intentional sensory complex. And this present resolution will consider only apparent nonlinearity, as mathematical nonlinearity will require the evolution of neural organizations that deal with the retention of state, a process a little farther down the road of evolution, and a subject somewhat later in this dialog.
Returning to our imagination scenario of the early evolution of the orientation stage in neural arrays, with a current focus now on the sensory complex, we can see how the emergence of the emotive complex will provide the molding forces for Natures’ adaptation of exteroceptive electro-chemical processes that signal various cues in the environment, for which the sensory complex can provide suitable translations signaling aversive conditions and attractive conditions, as well as the just as important translations signaling the cessation of any triggered condition.
The simplicity in the neural construction of the early phasic and tonic aggregates allowed Nature to organize various configurations of these circuits into assemblies which accepted the neural signals from the electro-chemical processes, and these “input” assemblies developed neural signaling to the emotive complex and the metabolic neural complex, representing the “sensation” and cessation of aversive and attractive cues in the environment.
And if we were to look closely at the exogenous development of the electro-chemical specializations themselves, we might see the differentiation of the molecular sensors into the various types that evolution would select as those providing the most survivability in the environment of that organic species. And in the early phase of the species’ orientation stage, we might also see the exo-layer of the projective pole in the species’ locomotive axis being peppered with various experimental assortments of these electro-chemical processes, all of which possessed axonal appendages to transmit the binding of molecular and mechanical events to the sensory complex “input” assembly of the organisms’ neural array.
At this level of sophistication in the species, we would observe organisms that exhibited behavior no more complex than a phobic response to aversive conditions and a simple gradient response to attractive conditions, because the kinesthetic programs released by the motoric complex cannot gauge any true direction for the environmental cues being signaled. In the case of the phobic response, the present sensory complex can only tell the motoric complex which direction not to take, and its gradient response to attractive conditions carries little more information.
So before Nature could begin the specialization of the exteroceptive processes into mechanisms that discriminate the “what” that is being signaled (besides the polarity of aversive and attractive gradients into various degrees), the “where” of these gradients was to become the next evolutionary priority. Here again, the motoric complex continues to drive the adaptation of the sensory complex.
Because of this new evolutionary emphasis on the sensation of directionality during the adaptation of our organisms, we would see a migration in the peppering of the electro-chemical processes of evolving organisms as they coalesced into two concentrations, which we shall term as “buds”, using the term in a very simple, functional sense, without the many layers of biological connotations it typically conveys. And for selective reasons that do not bear explanation, these two concentrations would migrate over the course of evolution to co-locations on either side of the projective pole of the organisms’ locomotive axis, still, of course, embedded at the surface of the organisms exolayer.
This new configuration in exteroception would enable a “tropotactic” response to environmental gradients, as opposed to the simple phobic response as before, by providing a second dimension to the sensation of environmental gradients. This second dimension would provide no additional information to discriminate the differentiation of gradients into “what”, beyond the overt specialization of the electro-chemical processes binding to specific molecules, but would provide the information to discriminate a relative direction for the environmental signals. (A true “where” discrimination will require the virtual construction of a third dimension by a “cognitive function”, which is very much giving away the plot, but again the dialog could not resist here.)
This tropotactic response, which in short order will evolve into tropic responses, will allow the motoric complexes in our evolving organisms to develop true taxes, or motoric sequences and their releasing mechanisms, which would permit the organism to orient itself in relation to a true angle between the environmental gradient and the locomotive axis.
But first, our evolving sensory complex would have to make “sense” of this new dimensionality, and so the bottom-up engineer should see that our imagination scenario has landed us squarely at the scene for which Nature must have been at to begin to solve the reconciliation of nonlinearity, because it should be remembered that the sensory complex has been so far constructed with phasic and tonic circuits, assemblies whose behavior is uni-dimensional, and whose response becomes nonlinear when exposed to the multi-dimensional, even this two dimensional, input of the environment.
Certainly Natures’ solution to this dilemma involved a great many experiments, but the dialog wants to emphasize how this phylogenetic response to apparent nonlinearity will begin the never-ending evolutionary climb up a ladder that neural arrays, and ultimately central nervous systems will take as they incrementally add response to the multi-fold dimensionality encountered in the environment. This multi-fold dimensionality of the environment will require multi-fold response mechanisms in the sensation complexes of evolving organisms, and this solution to the present dilemma comes as an adaptation within the primal sensory complex to respond to the contralateral signals coming from the evolved buds of dual uni-dimensional exteroceptors on each side of the locomotive axis, dual signals which spatially and simultaneously indicate different levels of the same environmental gradient, an adaptation which represents the primordial first step toward intelligence…
It is hoped that the meme of this primordial first step will also begin to tease out the beginnings of the formal concepts that the dialog has been alluding to in the mindset for the bottom-up engineer. As it turns out, the new neural circuit needed to develop this second dimension which could resolve a relative angular direction is conceptually straightforward, and it is conceivable that Nature did not have to perform too much experimentation to arrive at a solution that could be phylogenetically captured. And it is not hard to see how this solution would become ubiquitous throughout Natures’ diversity in organisms.
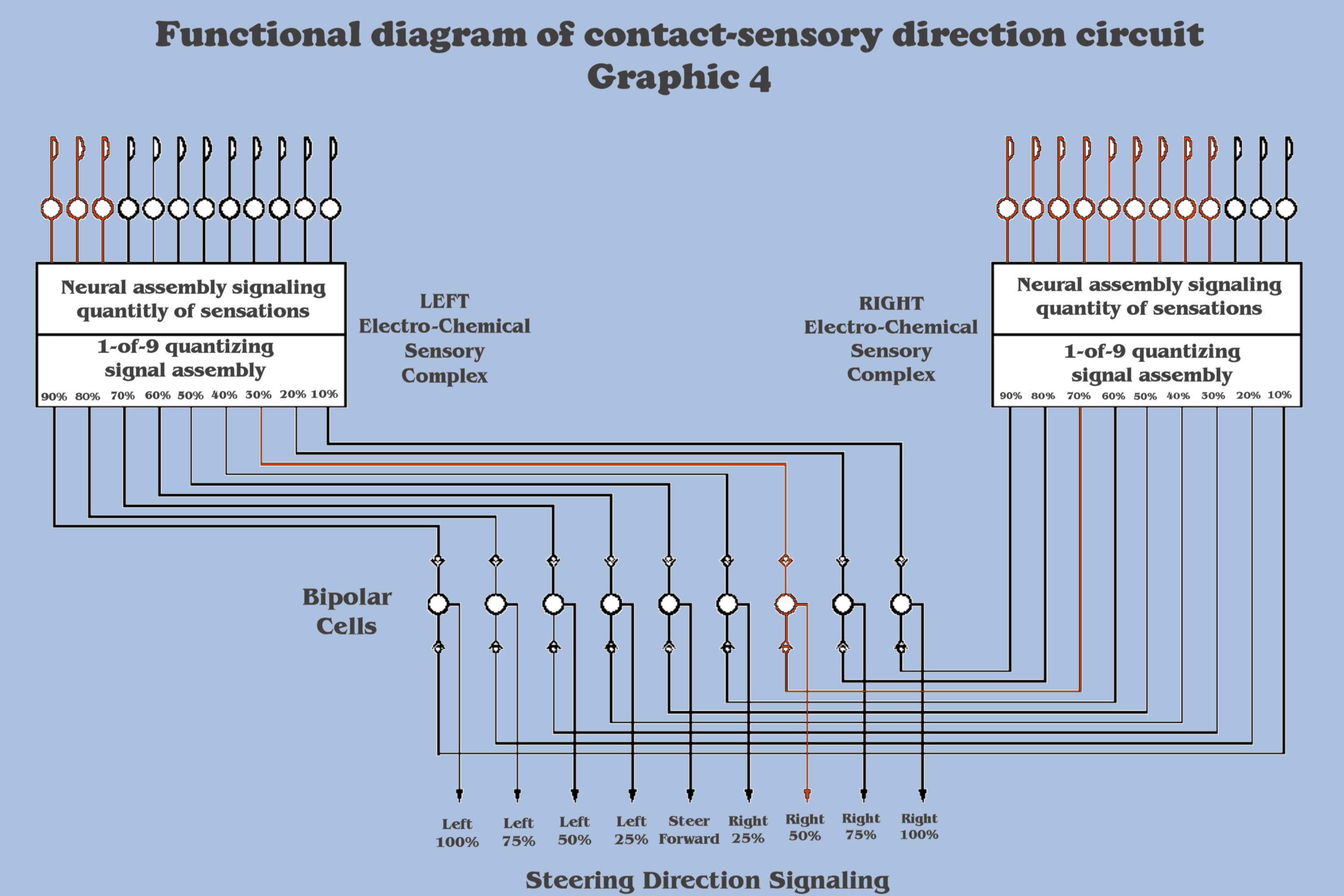
And again, the dialog needs to stress that this adaptation did not come about by the repurposing of existing components, a favorite technique employed by Nature, but by the specialization of a uniquely new neural assembly which represents the diffusion of two existing components. And the dialog also wants to emphasize to the bottom-up engineer how the systemic properties of this new neural arrangement is a direct result of the functionality provided by the development of the newly evolved relative angular “detector”, and how the bottom-up engineer should understand that this new systemic property contrasts with the more complex assemblies we shall soon encounter whose systemic properties arise solely from the structural interaction of their components, the very basis for the behavior of emergence that we seek.
At this point, the dialog could spend a whole chapter detailing the fascinating adaptations of the two coalesced buds of exteroceptors, as they evolve into the diverse forms of ramified antennae that have adorned many of Natures’ species, each form differentiated by the vagaries of specific niche environments. But there are so many more introductions on order and so little space for exposition that the dialog must remain focused on the development of the organic neural array, as we witness the progression of the orientation stage of the array into assemblies that can be called the pre-cognitive stage of neural complexity.

But before we return our focus to the neural array, there are two important aspects of the primitive exteroceptors that the dialog will return to in later discussions, aspects which should be placed in the back of the mind (another one of those puns) of the bottom-up engineer.
The first important aspect involves the specializations of the developing antennae in species as they evolve. The dialog just stressed that ramified antenna developed from the selective pressures which prioritized the “where” in environmental gradients over the discrimination of “what”. We will see in further discussions that the continued discrimination of “where” will require additional neural sophistications before the further development of antennae can contribute to this functionality. But evolution will not, of course, cease the still-important adaptations of ramified antennae, and as the dialog will later relate, electro-chemical sensation will ultimately form the neural bases for most organisms’ discrimination of “what” in the environment.
The second important aspect is in regards to the differentiation of the exteroceptors themselves. The dialog highlighted the migration of electro-chemical exteroceptors into concentrations, but it should be noted that mechanical exteroceptors would not follow this concentration, and indeed would follow an opposite migration, dispersing throughout the exolayer real estate, where selective pressures would ultimately site them at locations providing the most strategic sensation of mechanical events in the environment. This somatosensory sensation would in turn pressure those primitive exteroceptors that suffer the most mechanical perturbations to evolve into motive appendages as adjuncts to the locomotive axis. Again, the dialog will have much to detail for the bottom-up engineer regarding these evolutions.
Having mentioned those points, it is with some reluctance that the dialog must leave the fascinating adaptations of ramified antennae and mechanical exteroception, and fast forward our imagination scenario of the early evolution of neural arrays somewhat, past these fascinating adaptations as this second dimension discrimination ability of the primal neural array becomes ever more adept in its orientation functionality, and specializing the sensory complex into multiple, concurrent but independent “nodes” tasked with the second dimension processing of the chemically unique exteroceptive sensors that Nature will be perpetually experimenting with.
An experimentation that is always exploiting that same fundamental first step toward intelligence: the apprehension of simultaneous spatial differences in the same sensory idiom.
GO TO NEXT DISCUSSION
GO TO TOP OF DISCUSSION
Copyright © 2019 All rights reserved