As this exploitation of the newly developed second dimension allowed primitive organisms to graduate from phobic responses in the environment to true tropic behaviors, we would see those evolving organisms developing motoric abilities with increasing mobility and even more sophisticated behaviors.
However, this increasing mobility would have been brought about from a sensory complex based on electro-chemical receptors, a sensation medium requiring contact with the environmental gradient that it was signaling. And undoubtedly, this increasing mobility would allow Nature to evolve organisms possessing motive mechanisms that will literally outrun the perimetric sensory capabilities of contact exteroception.
Throughout discussion 9d, the dialog stressed the meme that it was the motoric complex which drove the early evolution of the primordial neural array, developing the emotive complex and shaping the sensory complex, all the while considering the metabolic complex as merely the coal car passively hooked to the steam engine pulling the train that was our primordial motile metazoan.
But now the metazoan train is beginning to outrun its meager lamplights. The increasingly mobile metazoan is developing behaviors that will allow it to blunder into environmental situations for which its tropic responses are increasingly unable to cope with. And the bottom-up engineer should by now recognize this as a now-familiar recipe for another evolutionary selective pressure. This limit on mobility that molecular and mechanical sensation placed on organisms would finally tip the balance of evolutionary adaptation toward changes in the sensory complex and the development of a different, non-contact transduction modality.
This dialog has characterized the development of the sensory complex as the climb up a ladder, the rungs of which represents the apprehension of successive orders of dimensionality in the experience of organic sensation. But the addition of each manifold stage of dimensionality was not always a gradual process. Sensory evolution also required epochal adaptations. And the first epochal sensory adaptation required the disconnection of primary sensation from electro-chemical molecular binding, and the adoption of alternative exteroceptors capable of signaling beyond the immediate surrounding chemical gradients of organisms.
And recall that even as the concentrations of exteroceptors were forming the first primitive antennae, Nature was experimenting with all manner of different neural receptors. The environmental signals being sent to the sensory complex did not have to be triggered by mechanical or molecular binding events alone. Even prior to evolving our motile metazoan, Nature had been experimenting with various forms of exteroception in the protozoans for many thousands of millennia, and she had already discovered the optimum medium for non-contact perception. Photoreceptor proteins had already evolved in some single celled organisms, allowing primitive discrimination between day and night to aid in metabolic circadian rhythms, the phototaxic orientation between up and down and simple depth control in aquatic metazoa.
So it is not a big leap to see Nature deftly adding to the electro-chemical exteroception for the motile metazoan in our imagination scenario, by including some photoreceptors in with the molecular binding processes. And these photoreceptors would provide a very unique signaling event to the evolving sensory complex, a signaling event which would constitute the next step toward intelligence. Certainly a non-contact sensory modality, this unique signaling event was not the reception of light, but the signaling of the occlusion of light.
In the pre-Cambrian and Cambrian seas, ambient daytime light was omnipresent throughout the coastal shallows, although only blue light would penetrate below 200 meters. However, at any depth, the immediate survival value to develop primitive photonic sensation and its most basic neural complexes would not have been in the gathering of light to form images, but in the gathering of light to detect its occlusion, signaling the shadows of obstructions, and more importantly, predators and prey.
In its earliest form, these primitive photonic exteroceptors would resemble minute patches of small numbers of these developing photoreceptors embedded within an organisms’ assortment of electro-chemical receptors. And it would not take much neural sophistication on the part of our motile metazoan to integrate the signaling of a sudden disappearance in the normally omnipresent light by a patch of these photoreceptors. And surely the bottom-up engineer can see how natural selection would favor the motile species that first developed this ability, and how evolution would promote the adoption of more complex capabilities to discriminate objects which did not fully reflect, or in many cases, occluded altogether the daytime ambient light. Natures’ first exploitation of photoreceptive exteroception would come in the form of a triggering of shadows.
And so naturally, just as Nature initially evolved ramified antennae out of a priority to discriminate the “where” in environmental gradients over the “what”, selection pressures would evolve visual exteroception initially from a priority to better discriminate the “where” in this triggered darkness over the “what” that might come from the imaging of the light itself.
There is a significant amount of literature discussing how the lensed ocular organ developed in species, and it is not the intention of this dialog to add to that speculation, only to emphasize to the bottom-up engineer that the adaptations which resulted in visual sense organs in species evolved in stages, and those stages were demarcated by the subsequent neural sophistication that had to develop to service each succeeding stage of the sense organ itself.
And much has also been written regarding how the aforementioned patches of photoreceptors would grow somewhat, and, due to the selective pressures to further discriminate the directionality of shadows, would over generations form a cavity or pouch and eventually fold back on itself, ultimately closing in to form a narrowed light aperture. Here again, there is much literature on how crystalline lenses were formed, probably first as a transparent barrier to keep foreign matter from fouling the sensitive “pouch”, and in our imagination scenario, as we are envisioning the formation of the first proto-eye, the discussion here wants to highlight the selective pressures which would evolve the proto-eye into the primordial compound eye, the morphological form of ocular organs which began the sense of vision in the early Cambrian era.

Although, certainly, the collective arrays of compound eyes did not develop all at once, it might seem incongruous that the compound collective visual organ would evolve before the functionally different dual-lateral vertebrate eyes.
The compound eye developed first because the neural arrays of Cambrian period organisms had not developed to a sophistication which is necessary to support the unitary lens-retinal vertebrate eye structure and the imagery of light.
The bottom-up engineer should not be deceived by the appearance of optical functionality in the compound eye. It is not like a remote video camera wired to a recorder. To begin with, none of the individual orbs of the compound eye would have the neural or muscular support for ocular accommodation, which is a change in lens shape by ocular muscles coordinated by reflex nerves in the vertebrate brain, a process necessary to focus light for imagery. And even if primordial compound eyes had the ocular machinery to effect this complex process, effective ocular accommodation requires a separate optical mechanism to move each ocular orb independently and produce ocular fusion, giving the optical capability to fixate a light radial on the same retinal region of each lensed element, a process requiring even more neural sophistication, and a process that would be impossibly complex with compound eyes.
The individual lensed elements of arthropod compound eyes are called ommatidia, and each ommatidium contains a cluster of 6 to 9 photoreceptor cells surrounded by support cells. The outer surface is covered with a transparent cornea, below which is situated a cone-shaped crystalline element. The crystalline cone is not a true beam-focusing lens as we would find in cameras and vertebrate eyes, but a light collector.
The cone collects light onto the tip of a rhabdom, which is a thin, long light guide, acting to further enhance light collection. Arrayed around the light guide in a roughly hexagonal pattern is the photoreceptor cells.
With only six to nine photoreceptors, an ommatidium initially would not be organized for light imaging, and in the primordial compound eyes, these lensed elements would best be adapted for the signaling of shadows. An optical mechanism capable of presenting a distance accommodated and optically fused image to the neural array would not have evolved before the neural arrays of organisms had developed the sophistication to abstract that imagery. The simple elements of primordial compound eyes were optimized for light gathering for sure, but the much more complex construction of retinal resolution and light imaging would take a number of epochal adaptations in neural sophistication.
The beginnings of that neural sophistication would come from the adaptations to accommodate the primordial compound eye, but before the dialog continues with the phenomenal development of primordial ocular sensation, the conversation cannot over-emphasize to the bottom-up engineer a concept which was stated at the beginning of this discussion – the concept that photonic sensation will have to subordinate electro-chemical sensation and become the primary medium to steer the locomotive axis. The implications of this subordination of contact sensation cannot be overstated because the bottom-up engineer should recognize that nothing in the sensory input from the primordial photonic apparatus can discriminate between attractive and aversive stimuli, and thus cannot directly contribute to the motility strategies of the emotive complex.
The phylogenetic implications of this consideration are enormous, and will be taken up in later discussions, but the bottom-up engineer must keep this consideration in mind as the subsequent evolution of photonic sensation unfolds. Although the early development of contact sensation was driven by the selective pressures which favored discrimination of the “where” of gradients before adaptations that allowed the determination of “what” in the environment, at least electro-chemical exteroceptors had developed the discrimination between aversive and attractive conditions.
But the locomotive axis still needed a steering mechanism controlled by non-contact sensation. And the successive adaptations which evolved the primordial compound eye developed an increasing discrimination in the directionality of shadows, a directionality which brought an additional, unique component which is not present in the directionality obtained from the dual sensory sampling of primordial antennae.
In the case of the directionality afforded by electro-chemical antennae, the two sensory samples signaled by each individual antenna provided the singular dimension which was referenced to the organisms’ locomotive axis, allowing the resolution of a direction vector, and thus providing a steering angle for the sensed gradient relative to the locomotive axis.
With the neural signaling coming from the compound eye sensorium, the locomotive axis dimension was replaced with sensory signals carrying real information, as both the longitudinal and lateral axes were now definable by true sensory sampling, and ultimately, much like the dimensional resolution that was constructed in contact sensation, selective pressures evolved a neural array capable of the multi-dimensional resolution of a similar direction vector from the photonic signals, but in this case the vector was accompanied with a relative magnitude value. This new vector was signaling BOTH direction and distance. By contrast, the vector afforded by the neural machinery of contact sensation was just a simple unit vector – a general direction with no distance.
Our motile metazoan has scaled another rung in the ladder of dimensionality.
This additional piece of information was just what the organisms’ motoric complex needed to regulate the rate of the locomotive axis – full speed on the throttle when the direction vector was long, meaning shadows were distant, but a more measured movement when the vector was short, implying shadows were closer. Unfortunately, the remaining information in the direction vector had a fundamental flaw.
Although the information represented by the magnitude component of the direction vector could be taken advantage of immediately, the information represented by the direction component was not sufficient to control the motoric complex. Recall the imperative that before the motoric complex could be commanded to “steer left”, or “steer right”, or “steer forward”, the emotive decision to either “steer toward” or “steer away” from the shadow must be made. But unlike the organisms’ contact exteroception, the newly developing compound eyes could not provide any information to the emotive complex regarding the aversive or attractive polarity of these shadows. The information could give no guidance to steer away or toward the shadow. So until selective pressures could evolve a solution to this deficiency, the emotive complex would still be controlled by contact sensation, but with the addition of this new photonic sensory modality, the overall motoric complex was becoming increasingly schizophrenic.
This is a fundamental schism that many neuroscientists miss, probably because, in their defense, it is considered imprecise for scientists to think in such teleological terms. But the Organon Sutra has no such aversions to presenting these purposeful conceptualizations to the students of massively asynchronous assemblies and bottom-up engineers, even though evolution lacks any foresight. And had this increasing schizophrenia reduced the survival value of the developing photonic sensation apparatus, then metazoan species would not have seen any further evolution of this modality, but that was not the case.
The bottom-up engineer can see the phylogenetic implications of this fundamental disconnect when it becomes apparent that, although the photonic derivation network of the sensory complex is resolving a direction vector with a magnitude, only the magnitude component of the derivation will at first provide any survival value.
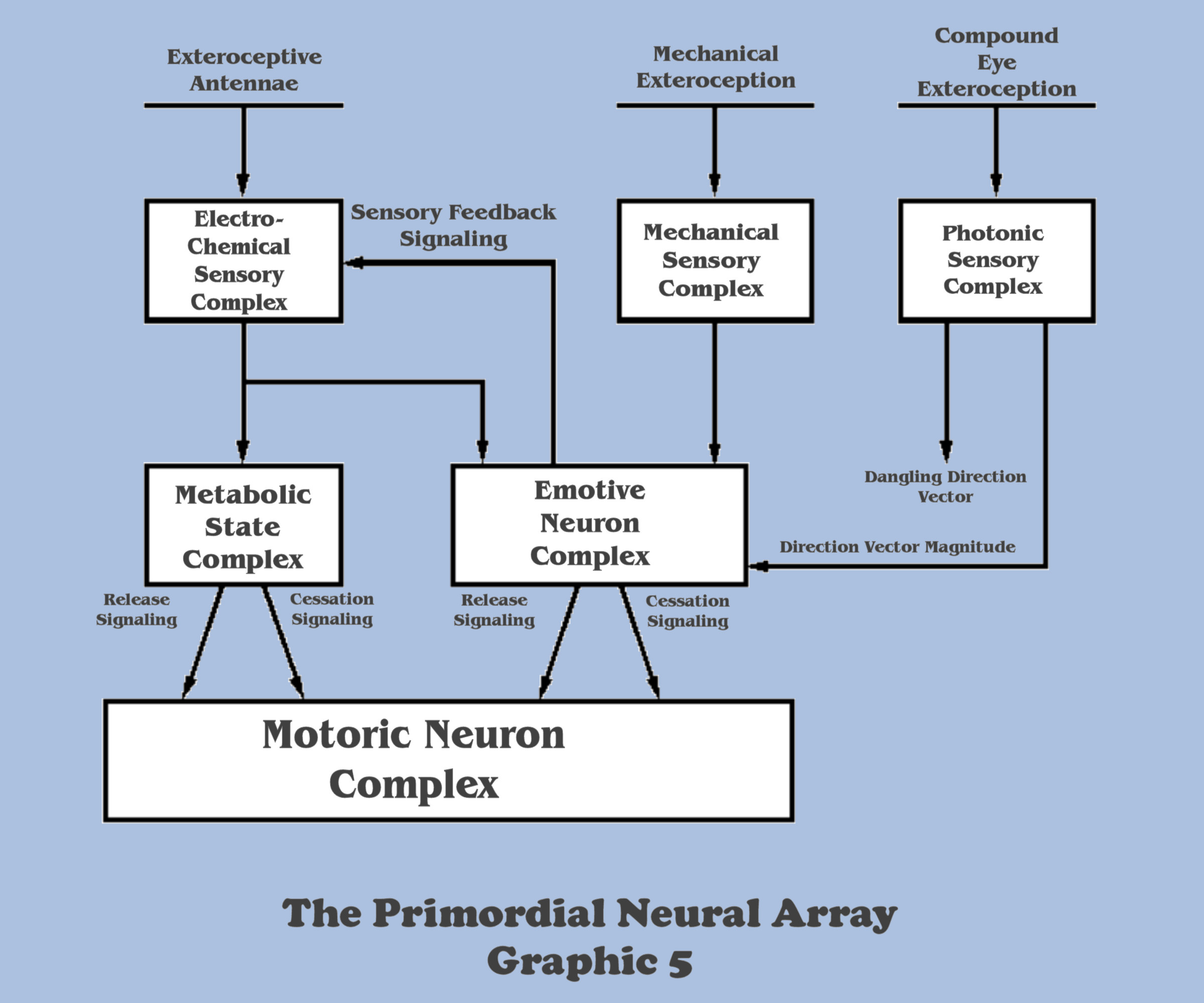
But the direction component of this derivation could hardly be just discarded. Nature is never so wasteful of her designs, and would never evolve an organ such as the compound eye, which demands so much metabolic resources of the organism for its ontogeny, simply to ignore a major component of its epochal contribution. And perhaps the abundant speciation displaying the evolving compound eye since the Cambrian era is indicative of Natures’ resolution to this disparity.
The additional dimensionality afforded by compound eyes came from the duplication of ommatidia and their physical organization of a grid pattern within the compound array. The portion of the sensory complex that is responding to the signals of the ommatidia are fundamentally organized into what electronic engineers would characterize as a summation network, and the derivation of directionality was still to be accomplished with essentially the same neural circuitry used for the derivation of the unit vector being signaled by contact exteroception. But the twist that Nature discovered was to duplicate the organizational geometry of the compound eye into the neural network providing its overall summation.
Now, the bottom-up engineer should not consider the signaling of each ommatidium to represent a pixel or individual picture element among many in a neural grid comprising a composite picture. The signaling from each lensed element represents a local direction vector indicating the directionality of shadows as sensed at that particular position of the compound eye, with each photoreceptor within the ommatidium acting like the heading point on a compass rose, (although because of its roughly hexagonal construction, this directionality was limited in its geometric resolution.) The growing sophistication of the primordial neural array had only evolved to the point of having the ability to sum all of the individual ommatidium direction vectors into a singular composite vector, but in this case, a vector also possessing a relative magnitude value. Although it is the resolution of a composite image that a central nervous system would evolve to, many more steps and epochal adaptations in the neural array would be needed.
The next major step in that adaptation lies with the settlement of the dangling direction vector, a direction vector which only has survival value once the aversive or attractive character of the shadow it represents is determined. Since nothing in the exteroception of the vector can provide an emotive component to the emotive complex, this information must somehow be abstracted from the available vector signaling, a process that will require neural circuitry that is physiologically different than any of that expressed in our primordial metazoan so far, and a process that will require another epochal adaptation.
And certainly this new physiology will require complex adaptations, and this new selective pressure will serve to introduce the Second Fundamental Precept of the Organon Sutra.
For those heroic readers who have persevered with the dialog so far, more engagement will be required…
GO TO NEXT DISCUSSION
GO TO TOP OF DISCUSSION
Copyright © 2019 All rights reserved