The prior discussion concluded with a conceptual visualization of a virtual layer cake formed from the many ideas surrounding the multi-fold adaptations which cumulatively evolved the primordial vertebrate eye. And we were about to slice through that virtual cake with an imaginary knife, in an effort to gain a different perspective on the so far unrelated rational significance between the layers of that conceptual cake.
From a conceptual standpoint, the whole cake represents Natures’ early neural processes to image the instantaneous signaling arriving at the primordial vertebrate optic tectum from the retinal ganglion cells of the organisms’ eyes. To fully accomplish this imaging process, the optic tectum would need to derive local cues from the “pop-out” image fields comprising the instantaneous signaling of the optokinetic process. But the bottom-up engineer will recall that the early optic tectum of the primordial vertebrate is quite unsophisticated in the imaging process as yet, and Nature would need time to phylogenetically adopt those behaviors which can provide perceptual cue derivation of retinotopically local photonic signaling, time that the survival of our early vertebrate species will not allow.
So, using our imaginary knife, let’s cut out a slice of that conceptual cake and see if we can gain another perspective on how Nature would come up with survivable behaviors to detect the emotive character in the emerged fields requiring visual imaging.
Somewhat toward the middle of the prior discussion, the dialog related a meme which could frame that perspective. Recall that if the bottom-up engineer were to conceive of neural maps as the functional part of a “procedural language” for global definitions in an “imaging semantic”, then the procedural imperative to maintain topologic coherence in those neural maps would dictate the need for syntactic forms which maintain that coherence across subsequent steps of the imaging semantic.
So, looking at the soon-to-be evolved imaging process needed for the early vertebrate eye from the perspective of that meme, before Nature could objectify the individual constraints behind any optokinetically emerged fields (before Nature could evolve local cues for recognition), there would have to be a way to discriminate between emerged fields collectively. Or, in the words of the topologic meme, before Nature can develop local cognitive semantics, she must be able to develop global perception semantics.
As always, the harsh lessons of the environment are relentlessly demanding behaviors that have immediate survival value, so Nature must fall back on her toolbox of existing neurological assemblies for those perception semantics.
Continuing the meme of visual topologic semantics, a perception systematic will dictate the syntactic definitions for these processes. Recalling the simple example of spatial invariance presented in the previous discussion, where the instantaneous “pop-out” signaling of a possible predator was impressed at some variable spatial location on the neural map of a particular hemisphere of the optic tectum, let us see how Nature would develop the neurological processes that might result in a spatially invariant representation of that possible predator image being impressed on a secondary neural map, ready for the immediate neural determination of its emotive character as aversive or attractive to our primordial vertebrate.
To be sure, the bottom-up engineer should know that the effective process just described is technically considered to be a translationally invariant function. Semantically speaking, a true spatially invariant transfer function would not only realign the local center of the emerged field from the retinotopic map to the global center of the (newly contrived) secondary cue analysis neural map, but it would transfer the image in such a way that the local 2-dimensional extents of the image field as it appears in the retinotopic map would invariantly fill the 2-dimensional extents of the cue analysis map, a transform of spatial scaling in addition to translation. But Nature is still new to the semantics of global transformations, and there will be immediate survival advantages to proceeding with just translational invariance from the start. And, of course, true spatial invariance would require the functionality to spatially scale the retinotopic image to an absolute scale, an operation that will call for cognitive abilities on the part of the neural assembly (oops, another plot spoiler…)
Since our primordial vertebrate requires an immediate solution to determining the emotive character of the emerged image fields developed by the optokinetic process, and especially the aversive character of predators, (detecting the aversive character of obstacles to locomotion in the visual field will be discussed later in the dialog), using a simple implementation of translational invariance combined with a mechanism which detects any motion of an emerged image field toward the organism would provide a tried and true trigger for an aversive avoidance reaction.
“Tried and true” in that this is the same avoidance mechanism developed for the compound eye “visual” system in Cambrian metazoans. And the methodology that Nature used to implement that Cambrian behavior involved principally metabotropic processes, which have since then been refined and are seemingly ready-made for a behavioral implementation in the evolving primordial vertebrate. The optokinetic mechanism has provided our vertebrate with a method to discriminate the motion of potential predators relative to the surrounding background of the organism, and now Nature will rediscover the survival value in evolving a mechanism to discriminate the motion of a potential predator relative to the organism itself.
Perhaps it would be a straightforward process for the optic tectum in our vertebrate to develop the capability, using a newly evolved transform semantic of global invariance, to transfer the emerged “pop-out” image field from the primary retinotopic map to a secondary cue analysis map in a translationally invariant manner, centering the image field in the secondary map, and thence, metabotropically abstracting the temporal state change in the field with subsequent temporal states of the image, to determine if the movement of the image field is aversely toward the organism or not.
Metabotropically abstracting the change in the instantaneous signaling of the sensory apparatus is a behavior which Nature has successfully engineered before, but how would she go about evolving this new transform semantic of global invariances? Surely Nature would implore the dimensionally localizing qualities of metabotropic state synthesis, but the adaptations to abstract map-wide invariances in the neural maps of the optic tectum would require the combination of both temporal and spatial state synthesis, a new metabotropic mechanism that can differentiate temporally disjoint signaling occurring over spatially disjoint synapses.
From an engineering standpoint, Nature has encountered spatial state synthesis already, in the operations performed for contrast extraction, which is essentially an abstraction of the state in comparison of illumination levels between spatially adjacent light receptors. And we have seen how this was accomplished both metabotropically and mechanically. But we have also seen how contrast extraction already has a temporal component to it, which is why it could not be implemented merely with a neurally connected phasic ensemble. So, although engineering spatial state synthesis by itself is already proven, combining this process with another metabotropic process having an entirely unrelated temporal component will require new neural behaviors.
This adaptation process to develop map-wide state synthesis behaviors in the primordial vertebrate neural array is being presented from a very teleological perspective to assist the bottom-up engineer in envisioning the neurophysiological dilemma that Nature faced in evolving an abstraction mechanism that combines temporal state synthesis with spatial state synthesis. Natural evolutionary processes act on the system as a whole, not on its parts, but the bottom-up engineer must approach the engineering of her systems from the bottom up, by contemplating the parts before the system (arguably very difficult to do!)
To help with this, the bottom-up engineer is reminded of the dialog in discussion 10B, where the basic nature of the neural metabotropic mechanism was illustrated. Recall that in the discussion, ionotropic persistence could occur at any synapse, but dendritic charge accumulation from these persistences, (the collective charge depolarizations that could lead to firing of an axonic action potential), occurred only between spatially localized synapses in the dendrite.
The metabotropic behavior that is needed for map-wide state abstraction requires a mechanism to produce ionotropic persistence at synapses which may not be spatially localized at the dendrite. Nature will have to get creative again with neurophysiology. Spatially distributed ionotropic persistence is a behavior which is moving closer to the classic definition of memory.
Returning to discussion 10B, metabotropic persistence was presented as the result of a spatially localized release of calcium ions from the smooth endoplasmic reticulum, which significantly delayed the electrotonic dispersal of ions gated into the dendrite from synaptic activity at that spatial location.
But the new metabotropic mechanism for spatial state synthesis needs to behave in a different manner. Instead of producing temporal persistences in dendrites, this new synthesis is more akin to determining the spatial pattern of synaptic activity as it is distributed throughout the entire dendrite over a certain time period. So the problem can be defined from another perspective: How can individual synapses change their behavior based on the spatial distribution of synaptic activity around them, even activity that might be spatially distant on the dendrite?
Variable synaptic behavior, or synaptic plasticity, has been the subject of intense neuroscientific research ever since the discovery of chemical messaging itself. But the majority of this research has been directed at the effects of activity at individual synapses in mammalian cortex. And this research is only reflective of extant organisms. Neuroscientists can only speculate on how much the mammalian neurophysiology has evolved since the neural adaptations that were present for the free-swimming vertebrate in the post-Cambrian period. So let us look at how Nature might have created the neurophysiological mechanisms of spatial state synthesis as we continue the imagination scenario of our evolving vertebrate in the Devonian period.
The bottom-up engineer should recall that the smooth endoplasmic reticulum (SER, also introduced in discussion 10B) in neurons possessed a number of qualities, one of which was the ability to selectively store and release much of the calcium ions inducted into the dendrite medium following synaptic activity. And in its role during the metabotropic processes of signal persistences, it acted as a mechanism to modulate and localize the diffusion of calcium ions within the dendrite. The SER undoubtedly started as a mere lipid and protein transporter in neurons, but Nature is always repurposing her designs, and in further adaptations, Nature gave the SER in neuron dendrites two other very important characteristics. The first characteristic involves the SER morphology. When it was first introduced in the dialog, the endoplasmic reticulum was called a “membrane within a membrane”, as it was an organelle that stretched from dendrite to soma to neural axon, and Nature has evolved the overall 3-dimensional topology of the organelle within the dendrites to be highly dynamic, with a morphology that can change over the time course of hours, ebbing and flowing within the circadian rhythm set up by the neural center of the metabotropic Locus Coeruleus.
The second important characteristic is that various regions of SER within the dendrites do not function equally in the storage and redistribution of calcium ions. The dynamic morphology of SER allows a localized accumulation and release of calcium ions which is modulated by the pattern of local synaptic activity, giving SER the ability to act as a sort of memory for extracellular events. The endomembrane system is the neuron behind the neuron. Where the ionotropic membrane mediates immediate signal thresholding, the endomembrane mediates a sequestration of signal history.
And how these characteristics are implemented to bring about spatial state synthesis is (superficially) illustrated in Graphic 14. In a very simplified and localized visualization of this sequestration, Panel 1 of Graphic 14 shows four spatially localized synapses, with a train of action potentials traveling along two of the pre-synaptic axons, and a singular action potential traveling along the second axon from the top.
In Panel 2 of Graphic 14, all of the action potentials have arrived at their synapses, and the induction and diffusion of calcium ions resulting from that signaling has produced charge fields similar to those seen in Graphic 9.
However, in Panel 3 of Graphic 14, we see something different. After an unspecified time period, the morphology of the SER depicted in the frame has changed, with filaments from the previously isomorphic organelle moving toward those specific synapses having the most pre-synaptic activity.
In Panel 4, there are illustrated two pre-synaptic axons with trains of action potentials approaching their synapses, and Panel 5 illustrates the resulting charge field from that activity, activity having a different synaptic spatial pattern that the activity which promoted the tubule encroachment illustrated in Panel 3.
In Panel 6, we again see trains of action potentials approaching their synapses, with a pattern in this case more similar to the pattern of activity exhibited in Panel 1, and in Panel 7 the subsequent charge fields resulting from that activity.
In Panel 8, the bottom-up engineer should notice that, although the morphology of the SER has not changed since the previous panel, the calcium ion concentration within it has changed, as it has absorbed the calcium ions produced from the previous volley of action potentials, whose charge fields were conformal to the SER morphology developed in Panel 3, a conformation which is reflective of the distributed spatial patterning of the signal activity demonstrated in Panels 1 and 2.
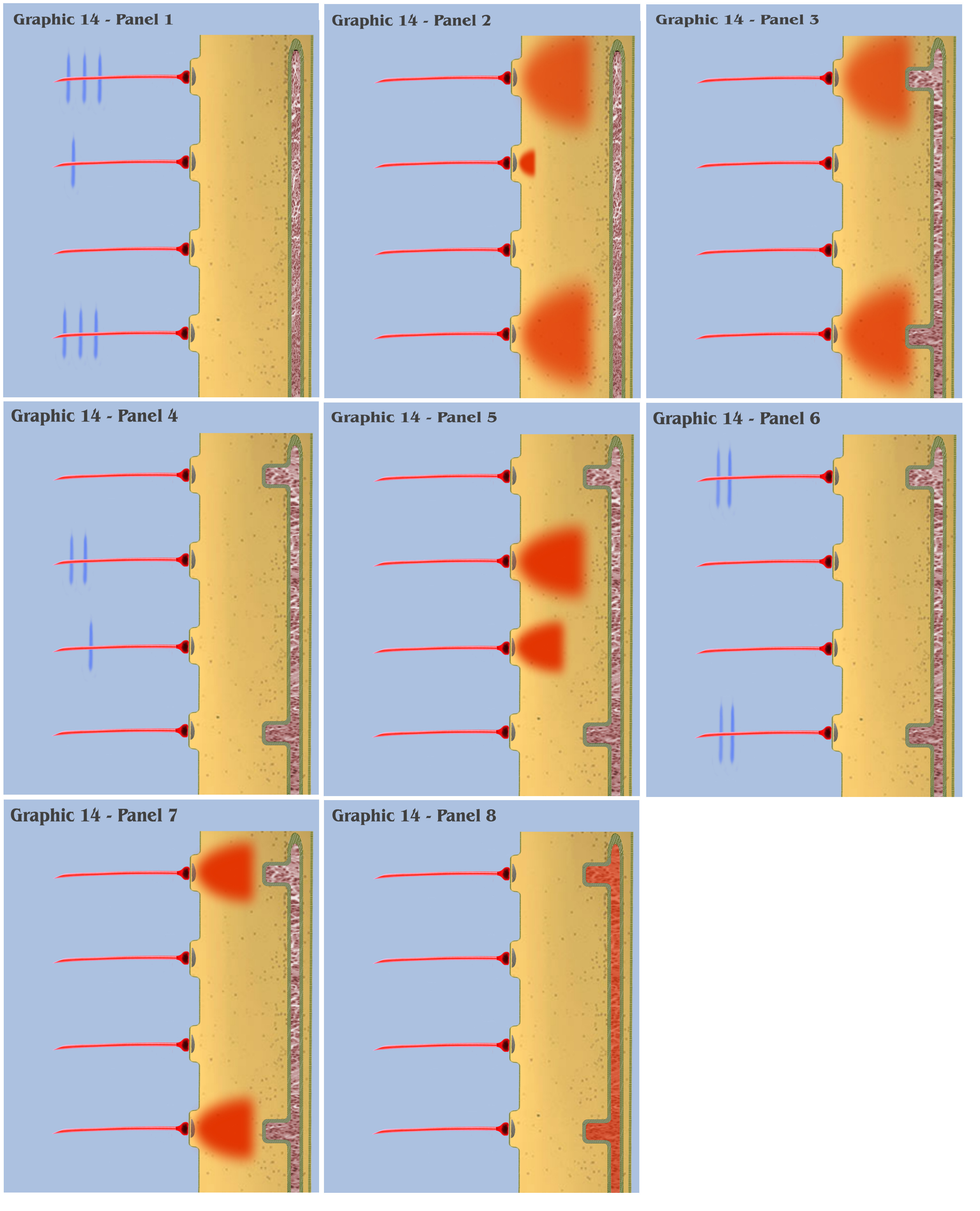
And the dialog wants to point out an even more important aspect of this visualization, as the bottom-up engineer should notice that the SER in Graphic 14 is depicted as open ended at the bottom. This is significant, in that it represents a continuation of the SER organelle, which was defined as continuous throughout the neuron, from dendrite to soma to axon. It is significant because of the final, important aspect of this new neuron physiology. In traditional neuron behavior, it is the accumulated charge at the dendritic membrane that is communicated to the neurons’ axon hillock, which, if it rises to the charge threshold level, triggers the ionotropically explosive process of electrogenic charge transmission along the axon. However, Nature has engineered a different process to communicate the triggering of the electrogenic charge parade for this new type of neural cell. In this new neurophysiology, an action potential is not triggered by the sum accumulation of charge potentials along the dendritic membrane, but instead by a repeat of pre-synaptic activity having a similar spatial patterning to a conformation of the dendritic SER (as depicted in Panel 7), whose dynamic morphology has been “molded” by previous patterns of synaptic activity (as depicted in panels 1-3). Because the body of the smooth endoplasmic reticulum is continuous throughout the dendrite, the activity which shapes its patterned morphology can occur at synapses with no local spatial relation to each other. And recall from Discussion 10B, where it was mentioned that the accumulation of calcium ions throughout the SER could serve as an intracellular messenger, and because the SER is contiguous within the dendrite and soma of the neuron, the endomembrane can communicate the “matching” pattern of synaptic activity directly to the cells’ axon hillock, creating a trigger mechanism for an action potential having a modality much different than the dendritic thresholding variety of neurons.
This new neurophysiology is beginning to cross the line between structures that demonstrate the elementary abstraction of state, and the true retention of state in organisms, getting us closer to a neural definition of classic “memory”. And perhaps soon, in today’s medical research, the key to a clearer neurophysiological picture of the memory debilitation symptoms exhibited at the onset of Dementia and Alzheimer’s disease will come with a fuller understanding of the long-term calcium ion sequestration properties in the smooth endoplasmic reticulum of neurons.
Combining this new neurophysiology, which in neural aggregates will express spatial state synthesis, with neural assemblies composed of the previously evolved metabotropic neurons expressing temporal state synthesis, provides Nature with the constructions that can collectively exhibit the emergent behavior of sensory perception, rising above the behavior of mere sensation in the exteroceptive apparatus, in a manner that could hardly be achieved with any hierarchical, connective approach.
These new constructions will provide the emotive complex in our early free-swimming vertebrate with the functionality to determine the aversive presence of predators, buying Nature the evolutionary time to phylogenetically adopt the remaining survival responses needed for approach behaviors to its prey and attractive mates. And it is during this phase that the neural assembly can establish the neural controls and refine the musculature for the final anatomical structure of the early vertebrate eye: the ocular lens which effects distance accommodation.
The organic mechanisms which effected the vestibulo-ocular reflex, along with the optokinetic response and intentional gaze fixation, were extraocular muscles that rotated the eyeball proper in its cranial socket. The muscles for distance accommodation, however, are located inside each eye, and their function is to move the lens in relation to the retinal surface inside the eye, and hence, change the focal distance of the eyes. And like the three functions just mentioned which alter the gaze angle of the eyes, the function of distance accommodation should really be considered within the context of the overall primordial imaging operation as a whole.
Extant fish and amphibians perform ocular accommodation by moving the lens relative to the retina, where birds and mammals use the more evolved process of deforming the lens, probably because the lens in fish is more spherical in shape, and there is little binocular feedback to control the lens positioning in fish while the eyes operate in an essentially monocular fashion during the optokinetic process. The mechanical process by which fish focus their spherical lens is well documented, but the neural processes directing this accommodation has not been fully researched. In terrestrial tetrapods with binocular vision, distance accommodation of the optic lens is a straightforward reflex response to the binocular gaze divergence. As the binocular gaze of the eyes diverges, when the animal is focusing on a more distant object, there is a direct reflex response of the lens muscles to focus farther away.
And certainly the refinement of this process of distance accommodation and lens positioning in early vertebrates will play a large part in the phylogenetic adaptations that our organism will develop for the approach behaviors to its prey and attractive mates, along with the integration of non-visual signaling (such as olfactory, auditory and lateral line modalities), but all of these neural adaptations will ultimately follow from the “imaging semantic” required to maintain topologic coherence in the imaging process, developed to detect aversive predators, where it was thought that the imaging process would in time develop “cues” in the imagery that the environment presented to our organism. However, those “cues”, (in the classic definition of local image elements which elicit recognition), require the cognitive ability of retaining prior experiences of the world, a level of neural sophistication which our Devonian vertebrate, only now emerging from mere organic sensation to neural perception, does not yet possess.
And yet, in a misplaced desire to explain cognitive behaviors before there has been a fundamental understanding of neural perception, neuroscientists studying extant organisms often overlook the remarkable level of sophistication that primordial vertebrates must have expressed, demonstrated in the imaging processes so far evolved. What is overlooked is not the lack of cognitive capabilities, but the development of an imaging semantic possessing domain transformations such as global invariance which are independent of the constraints present in the vertebrates environment, independent of the classic “cues” which our primordial vertebrate cannot yet perceive, and can only develop phylogenetically. What is often overlooked is how these independent domain transformations will become an essential substrate to those cognitive behaviors yet to come.
Since classic Fourier transforms are uni-dimensional, they do not have a direct application to the manifold formalisms of massively asynchronous assemblies, and likewise are not the conceptual solution for Natures’ current problem in vision analysis. For the bottom-up engineer, what is required for both of these realms is more of a combination of the conceptualizations of invariance that Fourier’s equations bring to the discourse, combined with the non-Euclidean geometries of Tensor math (Tensor math having those non-intuitive constructions that can curve the very dimension lines within a space), all this wrapped up with a nod to dynamic topology. This combination, this “essential substrate”, which the Organon Sutra has termed “Topodemics”, (a mathematical approach, although very non-intuitive to the human intellect, due to its temporal properties, which is perfectly suited to the hyper-dimensional specializations of artificial agents, and is considered to be a symbolic basis for dynamic tensor invariance, where all definitions use parametric methods to frame invariants), which will provide the bottom-up engineer with a beginning foundation for building the constructions that will express behavioral cognition in their artificial agents, solid developmental foundations for which hierarchical design methodologies can only theorize about.
Because the path to emergent intelligence can only be found with a resolution to the contradictory state which comes about when we try to define invariance within dynamic, fluid topologies, it will be instructive to see how Nature resolved this contradiction, a resolution that will lead the bottom-up engineer to the doorsteps of gestalt abstraction itself.
After that plot tease, let us wrap up the current discussion, because, like the fascinating developments which evolved ramified antennae in our primordial metazoan, the Organon Sutra could devote entire chapters to the remaining adaptations of the primordial vertebrate, as the various species establish their position in the evolutionary progression toward extant species today. But the compelling developments which will evolve cognition in organisms are beckoning, and so the dialog will close this discussion with the final behavior of the primordial free-swimming vertebrates that deserves mention.
At the beginning of this discussion, the dialog promised to cover the last aversive characteristic that pelagic organisms needed to develop for survival before any phylogenetically evolved approach behaviors: the detection of obstacles to locomotion in the visual field.
Recall that the optokinetic process was synchronized with the rhythmic phases in the fin-strokes of the fish, and just preceding the process that resulted in the emergence of the “pop-out” image fields from visual gaze fixation, the gaze angle of both eyes were shifted to a forward looking position in a saccadic action at the beginning of the fin-stroke cycle. At this point, even though the gaze of the eyes would be momentarily fixated, nothing in the visual field would “disappear” from the ganglionic contrast extraction process because everything in the visual field would be moving.
And within the overall visual field of both eyes, there would be a narrow band in the front, perhaps no more than 20 degrees, where the individual fields of view in each eye would overlap, promoting neural assemblies having increased sensitivity to anything in this field, a sensitivity which would represent an obstacle to the organism’s locomotion. This increased sensitivity will have pronounced survival value for the organism (species), and would not require much experimentation on the part of Nature to develop
Because of the pronounced survival value that this increased sensitivity will provide to the organism (species), the neural array within the primordial vertebrate of our imagination scenario will undergo some subtle adaptations. In Graphic 13, the optic fibers from each vertebrate eye serving the individual hemispheres of the optic tectum were portrayed as crossing over, with the optic tract from the right eye crossing over to stimulate the left hemisphere, and vice-versa for the left eye. However, because of the selective pressures brought about from the sensitivity to the field of view which overlaps in each eye, a small number of ganglionic axons comprising this overlapping field would not cross over, but will terminate in the optic tectum on the same side.
This adaptive arrangement will result in a small population of neurons in both hemispheres having co-incident stimulation from both eyes. And with this co-incident stimulation, it would not take Nature long to create simple phasic circuits within this small population of neurons that would register the phase coincidence of the signaling coming from separate eyes having overlapping, but identical, image fields. And this “phase-lock” circuitry would represent the very first non-metabotropic temporal signaling created in our evolving neural array. For sure, the basis for this temporality lies not in the connections of the phase coincidence neurons, but in the mechanical arrangement of overlapping signals from separate eyes. However, the temporality still represents Natures’ first step, and a most important one, toward non-metabotropic neural states, a first step which will set in motion phylogenetic changes having evolutionary ramifications hundreds of millions of years later.
Although this phase-coincidence circuitry might suggest a binocular imaging capability, the sophistication of the neural array in our primordial aquatic vertebrate at this stage has not evolved to a degree which could promote binocular perception, but this newest neural behavior will allow the dialog to close the current discussion and usher in the Third Fundamental Precept, which has been alluded to so many times.
GO TO NEXT DISCUSSION
GO TO TOP OF DISCUSSION
Copyright © 2019 All rights reserved