Since complex biological systems cannot process the entirety of the immediate environmental signaling being sensed at any given moment, Nature has found that there is survival value in being attentive to certain aspects in an organisms’ sensation while (safely) ignoring others. This discussion is intended to convey to the bottom-up engineer that there are many critical components to this process of selective attention, a mechanism which forms the first processes for the higher level behavior of self-organization, which is itself a paradigm for the processes of all biological adaptation.
As it was first introduced toward the end of discussion 9D, the paradigm of perceptual diffraction is meant to refer to one of the biological processes that Nature has evolved to bring about this capability for selective attention. Somewhat like a prism that refracts white light into its constituent spectral colors, wave diffraction is an effect which makes certain waves appear to travel in different directions after encountering an obstruction in their open medium. And just as the physics of wave diffraction cause physical waves to appear to travel differently, the organics of perceptual diffraction make the sensations of an organism appear differently in its neural array, giving an organism the ability to attend to different aspects of the environment, depending on whether the organism is in an aversive environment or an attractive environment. As a form of “selective resolution”, perceptual diffraction is the mechanics whereby an organism can make those environmental cues that are relevant to a particular aversive or attractive environment “come into appearance”, allowing the organism to develop those response behaviors having the most survival value.
After that specification, the bottom-up engineer might first ask by what process it is that the relevant cues in a particular environment are chosen for attention in the first place, but before answering that, the dialog will initially consider the actual neurophysiological mechanisms that can promote some signaling of sensation while attenuating others.
Because of the reality that the environment might present dangers to an organism as well as attractions, the dialog has stressed the meme that mobile organisms cannot afford to be just passive occupants in the environment that they inhabit, survival demands that they become an active agent in that habitation. As this selective pressure has been the genesis for many adaptations, including intentional mobility in the evolution of organisms, the bottom-up engineer should also expect to see adaptations leading to the intentional sensation of the environment. Since mobility can no longer be performed in a passive manner, certainly sensation of the environment can no longer be a passive activity.
The paradigm of organic sensation as an intentional activity has been declared as one of the central tenets of the Organon Sutra in discussion 9D, where it was mentioned that this functionality was brought about by a feedback mechanism between the primordial metazoans’ emotive complex and its sensory complex. In that discussion, the feedback mechanism (illustrated in Graphic 3) was necessitated by the probability that different environmental cues would signal the detection and subsequent extinction of an aversive environment condition, allowing the emotive complex to direct the onset and cessation of mobility commands which would mitigate the aversive condition. But the evolving photonic sensation apparatus disrupted that simple scheme, (because photonic sensation cannot develop the cues needed to signal an aversive condition as a matter of its sensation), and this prompted the evolutionary adaptations of the primordial metabotropic processes, and the ability of the neural array to abstract state in its sensation as a substitute for the missing emotive signaling.
Now the emotive complex has a new responsibility, that of synthetically abstracting the emotive character of the signaling provided by the photonic sensory apparatus, and this new functionality will certainly change the feedback between it and the sensory complex, changes contributing to adaptations that will evolve the entire organisms’ neural array into the organizational phase that the Organon Sutra will characterize as the pre-cognitive phase, being the successor phase to the species’ orientation stage.
The first change to the feedback channel between the emotive complex and the sensory complex will come about from another physiological imperative of the recent adaptations in our motile metazoan. The persistence mechanism in the newly evolving metabotropic neural assemblies must not operate continuously, and must operate only during controlled periods, otherwise there could be no demarcation of individual states being abstracted by the mechanism.
It is the adaptations to develop this controlling mechanism that will result in the next fundamental re-organization of the motile metazoans’ entire neural array, a re-organization of a similar magnitude to the emergence of the emotive complex itself.
Continuing the imagination scenario of our evolving motile metazoan (species), the new responsibility of its emotive complex to abstract a synthetic discrimination between aversive and attractive conditions in the signaling of the photonic apparatus will result in a tri-lateral segregation of the organisms contact, electro-chemical sensory apparatus from its non-contact, photonic sensory apparatus, and its growing emotive complex. This tri-lateralization results from the neurological imperative that the previously singular feedback channel between the emotive complex and the sensory complex as a whole must now evolve into two feedback channels, each carrying disparate signaling, with one channel between the emotive complex and the sensory complex that is processing electro-chemical antennae exteroception, and a separate channel between the emotive complex and the non-contact sensory complex that is processing the photonic apparatus.
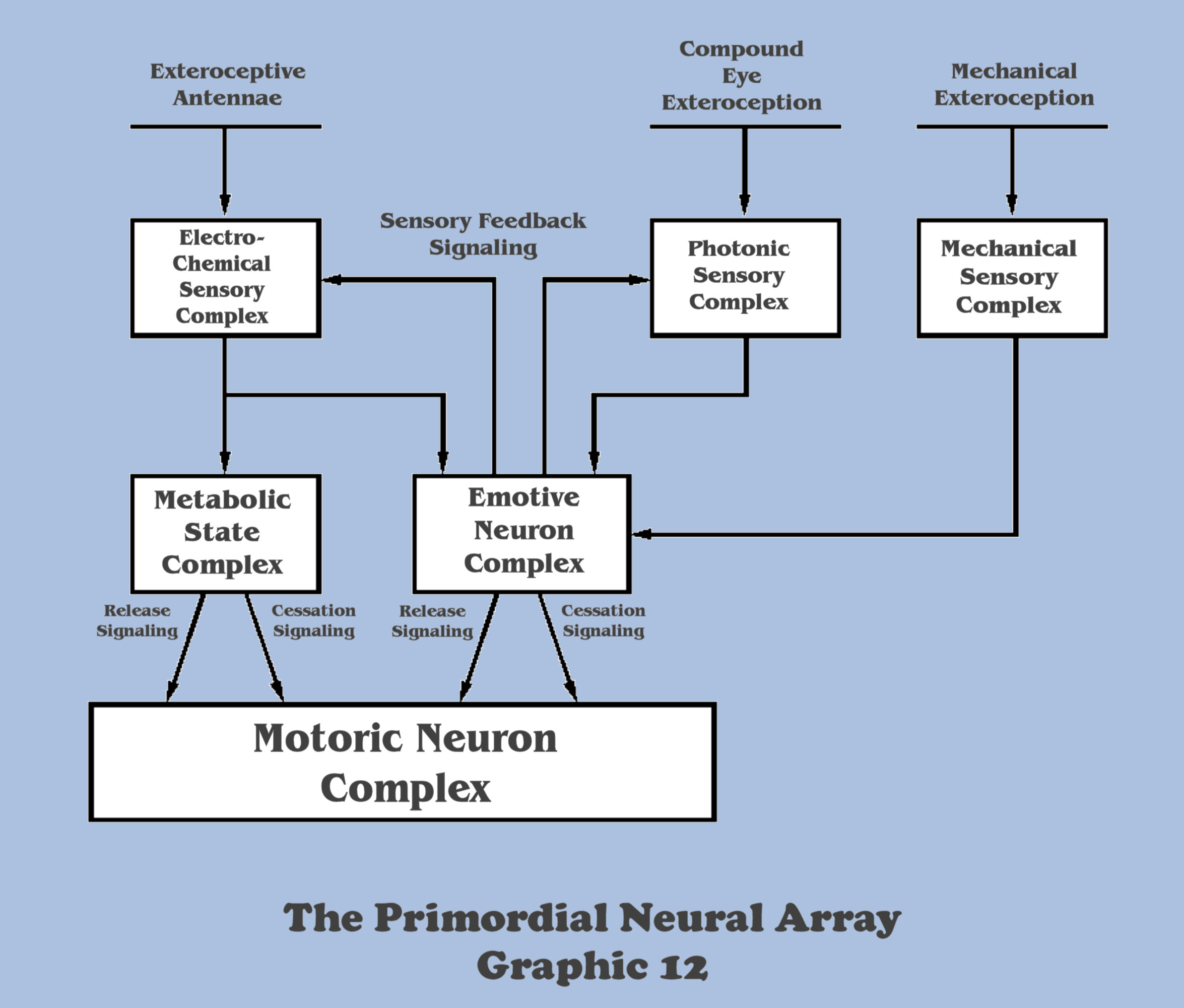
The bottom-up engineer can understand the distinction between the two feedback channels when it is pointed out that the electro-chemical exteroceptive apparatus of the organisms’ ramified antennae already provides the emotive cues to develop a discrimination between aversive/attractive conditions (in addition to the direction vector it develops for gradient concentrations), and does not require metabotropic feedback for its synthetic determination. The photonic sensation apparatus on the other hand, while providing a similar direction vector to its sensory idiom, cannot organically determine the emotive character of its signaling, and requires the controlled feedback of metabotropic biasing to synthesize the emotive characterization.
It is the development of the controlling mechanism in this feedback that is the focus of this discussion because the dialog wants to caution the bottom-up engineer from forming any pre-conceived notions regarding the “context basis” this mechanism might utilize for establishing its logical control “decisions”, pre-conceived notions that typically accompany the deprecated hierarchical thinking of a “controlling mechanism” which asserts its controlling behavior through a pre-established set of non-contradictory logical decisions.
While continuing its role as the stern, boot camp drill instructor, the Organon Sutra must diligently exercise the hierarchical mindset out of all students of massively asynchronous assemblies by instilling a strict discipline of bottom-up engineering, and in this case we want to be careful how we characterize the “control” mechanisms for this feedback channel between the emotive complex and the photonic sensory complex. Because this feedback is modulating the temporality introduced into the instantaneous signaling occurring in the photonic sensory complex, the “logic” in the controlling mechanism of this feedback must itself be an abstracted state of the signaling in the emotive complex, and not a concrete, pre-established pattern of signaling that would be hierarchically engineered.
Now, the dialog recognizes that this definition is still a bit too fuzzy for the bottom-up engineer to comprehend at first blush. The actual “context” which this controlling mechanism will base its “logic” on to form its control directives must be a dynamic set of first and second order state abstractions themselves. But before the dialog can define what processes shape those shifting states in the first place, the discussion will have to point out that even Nature herself did not get a chance to fully answer that question at first.
To envision what happened, the dialog will return to the imagination scenario of our evolving motile metazoan (species), at the point in its development where the tri-lateralization of the sensory complexes and the emotive complex has just occurred, and where Nature would begin to establish those adaptations that would begin to define the controlling characteristics of the feedback channel between the emotive complex and the photonic sensory apparatus. Although the motile metazoan (species) currently processes the entirety of its immediate sensory environment, the emerging behaviors of perceptual diffraction and selective attention will soon have the same immediacy characteristic of all of the selective pressures that have shaped the organism so far.
We have just witnessed how Nature solved the dilemma of the dangling direction vector by developing metabotropic processes that allowed the epochal realizations of state abstraction and the creation of synthetic emotive components to photonic signaling. And to maintain a separation between the instantaneous signaling of the sensory complex and the temporally fluid signaling of the synthetic processes, the imagination scenario was just about to resolve the feedback mechanism being created between the two.
The bottom-up engineer can certainly imagine that evolution could develop this feedback mechanism into a variety of neural adaptations, but before Nature could explore much of the wide-ranging possibilities that this mechanism offered, that harsh taskmaster, the environment, would present Nature with the next lesson in nonlinearity, a lesson that would form the mold for the neural organization that our evolving metazoans will need in the next epochal step in neural sophistication.
This next evolutionary step will require other seismic adaptations, as the unrelenting lessons of the environment again prod Nature into developing a breakthrough in neural organization, one in which will result in the transition from the signaling of shadows to the imaging of light itself.
GO TO NEXT DISCUSSION
GO TO TOP OF DISCUSSION
Copyright © 2019 All rights reserved