Life is a nonlinear experience. And that experience, as it rises from the local level of immediate sensation, through the global level of objectification, and finally through the introspective level of cause and effect, remains a common thread in the ability to detect regularities, the invariants, among spatial and temporal nonlinearities, which is a key aspect to intelligent behavior.
As the geometric area of their habitation expanded and outpaced the developing cerebral cortex in our primordial mammals, selective pressures provided the need for expanded sophistication in the exploitation of the association maps being abstracted by the hippocampus. To this point, the retrosplenial cortex in smaller mammals had been successful in the spatial localization of salient allocentric invariants, those “landmarks” of the current environment assimilated into the animals’ day-to-day behavioral repertoire which provided survival advantage apart from the organisms’ phylogenetic adaptations. But the inexhaustible variety of an environment will never let Nature rest with her creations. Even within the lifetime of a single organism, many of the environmental proxies for which the animal develops “landmarks” would change, possibly negating the survival advantage of their assimilated allocentric invariance. This special memorization was now in need of some temporal sophistication, for which Nature would evolve what neurologists have come to call the pre-frontal cortex in central nervous systems.
This structure is called the pre-frontal cortex because, recall that a frontal cortex has already been developing just anterior to the cerebral triad of sensory modalities, a structure which became the cerebral counterpart to what this dialog had been characterizing as the “motoric complex” in primordial vertebrates. (The bottom-up engineer should bear in mind that the cerebral component of the motoric assimilation was not all inclusive, as the basal ganglia had been retained for its phylogenetic storehouse of emotive, or involuntary releasing mechanisms.) As the pre-frontal cortex (PFC) develops in species, the central nervous systems in mammalian species will expand their sophistication in the exploitation of association maps. However, unlike the cerebral cortex of the smaller mammals, which drove the formation of those maps, the PFC will develop to deal with changes in metric/limbic association maps.
Although many neuroscientists consider the circadian rhythm to be the biological “clock” in organisms, for stimulus-bound animals, biological time is neurologically demarcated by the emotive state changes in many mammalian species. The circadian rhythm can only provide a cycle of continuity, whereas emotive changes in an organism provide true transitions to demarcate the formation of episodic memory, and the passage of biological time.
Now, where this demarcation in biological time is useful to the hippocampus in its function of abstracting association maps, and to the cerebellum in its sequential decimation reinforcing episodic memories, and ultimately to the retrosplenial complex in its normalization of the allocentric invariants of association maps, all of these neural assemblies are indifferent to the type of emotive state transitions occurring in their operation, being sensitive only to absolute states. And certainly, the fundamental reason for episodic memory is a mechanism to attach a particular emotive quality of aversion or attraction to individual allocentric invariants. But this attachment does not carry any dimensionality or any indication of the form of the transition between states, and therefore cannot provide any context to a neural assembly that may build behaviors in an organism to react to changes in its environment. Now, for hundreds of millions of years, Nature has been getting crafty at creating synthetic dimensions to unveil the many constraints that the environment hides behind apparent nonlinearity, and in evolving the functionality of the PFC, Natures’ craftiness will be no different.
To get a glimpse of this functionality, the dialog will ask the bottom-up engineer to step back from the PFC briefly, and consider the architectural state of the entire central nervous system in mammalian species so far. As we look at the overall organization of massively asynchronous assemblies in mammalian systems, we can generalize three classes of neuronal architectures.
The first class comprises the ontogenetically rigid neural assemblies that Nature has fashioned for what has been termed the reptilian and limbic structures.
The second class comprises an organization composed of only a few neural types, which behave asynchronously at the unit level but are specialized for parallel processing at the assembly level, and this structure is seen principally in the cerebellum of vertebrates. Because of this parallel nature, local interactions within the structure have a less influential effect on the assemblies’ output compared with the overall activity and connectivity of its input.
The third class of neural architectural forms, which comprise the cerebral cortex structures, are specialized for processing inputs which may be distributed from across multiple assemblies in the first and second architectures, and even interactive processing among inter-cortical structures. This distributed processing employs an almost universal recurrent network organization, where local interactions play a more significant role in an assemblies’ output than the activity of its input.
In the local organization of the cerebellum, inputs are tightly coupled temporally with outputs, and processing is accomplished with the cerebellums’ nonlinear distribution of spatial coupling.
In the global organization of the cerebral cortex, inputs might be temporally or spatially uncoupled from cortical outputs, and in the PFC, Nature expressed a specialization where inputs are temporally and spatially uncoupled from outputs.
The “virtual coupling” in either time or space is accomplished at the neural level in the dendrites of the neurons comprising these assemblies. Certainly, a major component of the cerebellums’ functionality can be attributed to the extensive dendritic branching of that assemblies’ purkinje cells, but its nonlinear spatial coupling of inputs to outputs can only be accomplished metabotropically, in the metabotropic modulation of the ionotropic receptors, called AMPA receptors, distributed throughout the dendrites of purkinje cells.
And also, within the cerebral cortex, capable of both temporal and spatial uncoupling of inputs to outputs, this functionality must also involve metabotropic processes, as the smooth endoplasmic reticulum in the spines of pyramidal cells perform a balancing modulation between dual ionotropic receptors. (AMPA receptors and so-called NMDA receptors. In the case of NMDA receptors, the smooth endoplasmic reticulum within spines provides selected receptors with an intracellular charge increment to variously overcome their voltage-gated nature, metabotropically switching selected NMDA receptors from unresponsive to responsive to pre-synaptic glutamate.)
Because the architecture of the cerebral cortex provides for distributed processing across all three classes of overall CNS architectures, metabotropic abstractions in the cortex are eventually differentiated between intra-connected processes and interconnected processes, a differentiation which was alluded to back in discussion 9C, and a differentiation that provided cerebral cortex with the flexibility to implement a spatial uncoupling or temporal uncoupling of inputs to outputs wherever it was functionally required. And this flexibility was expressed in its most powerful form in the PFC, whose organization specialized in an implementation that uncoupled inputs from outputs both spatially and temporally.
And, of course, as the bottom-up engineer might have guessed by now, the PFC achieves this specialization with another implementation of neural maps, aided by phylogenetically refined metabotropic processes, although in this case, maps having a distinct dimension of time.
By the time the PFC made its appearance in the neuro-architecture of mammalian central nervous systems, Nature had become very adept at implementing neural maps in her designs. With the concept of neural maps expressing a domain semantic which preserves the topologic coherence in two-dimensional transformations, all Nature needs to “build” neural maps are two metric measurement scales which can be neurologically associated in an orthonormal manner. By this definition, Nature should certainly be able to fashion a map having some spatial metric on one of the “axes”, and an orthonormal axis expressing a metric with a purely temporal dimension.
The PFC establishes this new biological time metric by creating a synthetic dimension to the emotive state transitions in stimulus-bound biological time. Although not a metric providing a true ratio scale, it is nonetheless a signed analog of the “swings” in the emotive state between aversion behaviors and attractive behaviors being demonstrated in the animals day-to-day activity. Through metabotropic persistences, the PFC “marks” various stages of this cycle back and forth between aversion and attraction, defining a repeatable measurement metric for the creation of semantic maps, very much like the association maps created in the sensory triad by the hippocampus.
And there should be absolutely no surprise on the part of the bottom-up engineer that, just as the cerebellum was recruited to produce episodic memories from the association maps of the hippocampus, the PFC would recruit it to produce semantic memory from these semantic maps in much the same fashion.
Although at this point, the bottom-up engineer might be understandably quizzical about one aspect of this recruitment. This dialog has detailed the participation of the cerebellum in the complex production of seemingly limitless motor sequences, or “muscle-memory” from the egocentric maps of the sensory triad, in addition to the production of innumerable “episodic memories” from the association maps created by the hippocampus, and now this recruitment has the cerebellum producing another entire collection of “semantic memories” for the PFC. In Graphic 15, a pictorial of the four major areas of the human brain is shown, illustrating the comparative sizes of the cerebellum relative to the other three areas for which it is providing all of this neural support, and so the bottom-up engineer might wonder why the cerebellum has not dwarfed the three other areas in size after all of this neural responsibility.
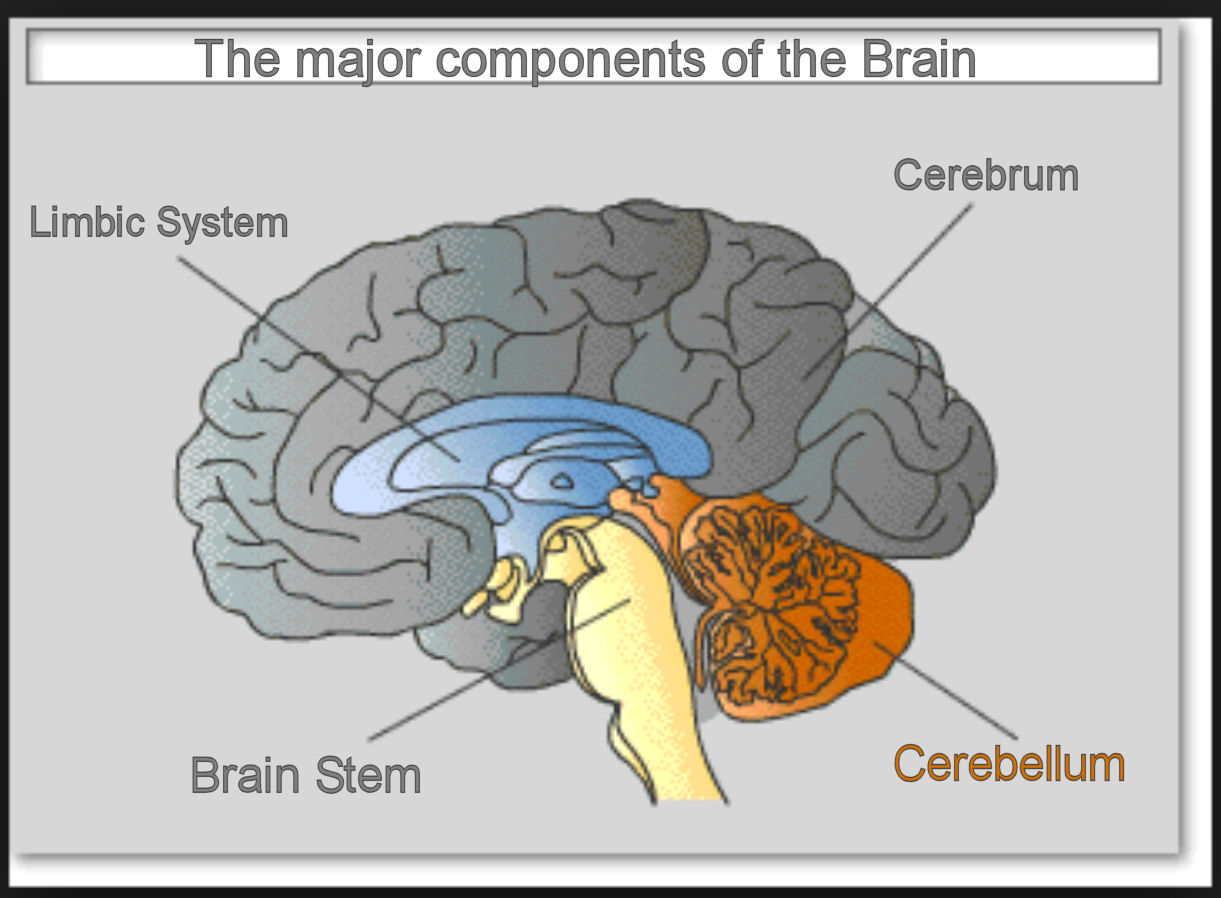
Although the cerebellum constitutes only 10% of the total weight in the primate and human brain, it is actually comprised of three times as many neurons as the entire rest of the brain. (Is it a coincidence that it is supporting three types of “memory”?) It occupies a much smaller volume because the majority of its neurons are much smaller granule cells, and this ratio has been conserved throughout the evolution of mammalian species. So, from a numerical standpoint, the cerebellum can grow in this functionality without crowding out the principle components of the brain.
Now, the dialog would like the bottom-up engineer to take a very specific perspective on these “semantic memories” being created by the cerebellum. It is not like they are a mechanism for the animal to remember what it had eaten the day before, (although it is a mechanism to “remember” where it found the path to that tasty colony of termites the day before), where the PFC is creating semantic maps, the cerebellum is establishing semantic spaces, and because semantic maps have a temporal dimension as an input, the successive “frames” produced by the cerebellums’ process of sequential decimation have a temporal dimension to this created “semantic space”.
For several hundred million years, Nature has been gaining experience in developing invariants in its maps, and now, in the PFC, it has learned to reverse the transformation of invariants, allowing the PFC to treat the semantic space as a pure temporal invariant, and abstracting away (uncoupling) the allocentric spatial component, revealing just the essential semantic “thing”.
Those “things” resolved by this new invariance transformation do not have a direct correspondence with a physical “thing” in an organisms environment, but these resolution products do represent an unchanging, specific aspect of the animals overall perception, temporal invariants in a ghostly “sense”, which are getting the organism closer to an ability to abstract the constraints which the wily environment has been hiding behind apparent nonlinearity ever since our primordial metazoan. However, there will be more epochal adaptations required before our primordial mammal can move from cognition to recognition.
GO TO NEXT DISCUSSION
GO TO TOP OF DISCUSSION
Copyright © 2019 All rights reserved