Since neurons are thresholding devices by nature, their triggering properties provide an ability to signal a summation of their inputs. However, Nature has exploited the different neural forms that have evolved to extend this summation property to a functionality that can perform either the integration or differentiation of its dendritic activity.
And Nature did not stop there. By further exploiting the morphological properties of neural membranes and the variety of fast neurotransmitters, Nature added the supplemental dimensions of spatial and temporal response, endowing some neural types with the capability to perform spatial integration, temporal integration, spatial differentiation and even a temporal differentiation response mechanism. Indeed, there are neural types which specialize in one or the other “processing modes”, and neural types that can combine multiple modes. Nature, it seems, rarely fails to exploit any variety that comes about from her own evolution.
And even that was not enough. In addition to these linear behaviors being fashioned into neural cells, Nature evolved other response modes, modes that do resemble computer logic devices in many ways. But to remain at arm’s length from the meme of “processing units”, the discourse on these tertiary behaviors must be delayed until the exposition of the remaining fundamental precepts, because, again, what those “bits of information” being integrated or differentiated are, is only relevant to the context of a particular pattern of activation in a neural coalition, the locus of processing, and the dialog has yet to fully develop the fundamentals of this tertiary behavior. “What” these neurons are differentiating and integrating should not be built into the devices themselves.
Just as we saw how Nature evolved multiple behaviors in singular elements by developing temporal as well as spatial integrators and differentiators, we would expect to see multiple behaviors being expressed in connected aggregates of individual units. It is these behaviors of aggregate collections that are the focus of the models being formalized for MAAs.
Returning to our model of natural intelligence, the least complex type of general activity in the neocortex is of a “phasic” type of collective response. This collective behavior of groups of neurons is a stimulus-bound state of activity, which is a type of activation that is dependent upon and persists only for the duration of some stimulus that has triggered the activity.
Phasic assemblies are characteristic in their response as they adapt rapidly to a stimulus, and phasic ensembles can be composed of a small or large collection of neurons, delineated by the extent or numerical distribution of the stimulus inputs to the ensemble. From the standpoint of individual neurons, phasic response to a stimulus also diminishes quickly and stops.
Phasic circuits are so called because their response typically follows some phase analog of their inputs.
This response does not convey information about the duration of the stimulus, but phasic neural assemblies are the predominant model for qualitative processing, especially in those cortical areas specialized for sensory processing. For example, in the cochlear structure of our hearing system, the sonic movements of fine hair cells are sensed by primary auditory neurons, whose phasic response represents some aspect of the several components of frequency and amplitude, components which are later integrated into “sound”.
Although typical of the phasic nature of processing, this response pattern of providing transformations upon a variable dimension of stimuli is not the only modus where we find phasic ensembles and coalitions. In many areas of the primary visual cortex for example, there are phasic assemblies that perform contrast enhancement of retinal input and assemblies for simple edge detection. There is no doubt that phasic ensembles can be created to transform just about any physical or abstract dimension of stimuli that natural selection finds advantageous.
The phasic type of neural assembly, typically organized around the level of a cerebral cortex mini-column or even whole columns (introduced in Discussion 9B), is a basic type of organization for not only the sensory processing areas of the neocortex, but is also found throughout areas specialized for motor control and general associative activity. Indeed, the neocortex is not the only brain structure which exhibits this form of assembly. The previously introduced limbic and “reptilian” brainstem structures have assemblies that demonstrate many phasic characteristics, although their functionality is more phylogenetically “hardwired”, and not as general purpose in nature as the assemblies found in the cerebral cortex.
The most distinctive aspect of phasic activity is that this model of “signal flow” is transformative, in that the phasic assembly receives stimulus activity in and responds with a linear transforming function out. And like we just mentioned with the limbic and reptilian structures created by Nature, whose transforming functions are fixed or dedicated phylogenetically, the ontogenesis of cortical columns and mini-columns provide them with a “programmable” quality allowing general purpose functional units, and we will delve deeper into this ontogenesis in later discussions. Now, looking at this processing from a hierarchical perspective, we would characterize this signal flow as unidirectional, but we want to be very careful with this. We will see in later discussions that the “direction of signal flow” is a very relative thing in the world of massively asynchronous assemblies. We will see that there is an almost universal expression of reciprocal connectivity in cerebral assemblies. That means that at any point in a neural formation, “direction” of neural activity can flow as before or completely reverse direction along reciprocal connections, and in most instances, simultaneous diverse patterns erupt into coalitions of neural activity.
The perspective that we do want to take is that of continually observing how one pattern of activity leads to another, irrespective of any arbitrarily imposed “direction”. The bottom-up engineer must always retain this “neo-think”. The science is in having a good characterization of what a pattern of activity is.
If we were to consider phasic activity to be the most basic activity in neural ensembles and coalitions, the next type of basic, general activity that occurs can be of a “tonic” type, which is an activity designed to maintain a balance, or a continuous maintained state. Tonic states are typically formed by establishing a closed feedback loop in the linear activation of neuron complexes. And Nature got very creative when she discovered this flexible and utilitarian form of assembly.
From a physiological standpoint, tonic activity is generally seen as a response to stimulus that is slow and is linearly graded. From a neurological point of view, tonic activation can be seen as neural complexes whose output is a linear adaptation to its stimulus. Because Nature found the need for a built-in response delay in tonic assemblies, this activation differs from phasic activity in that the response is typically measured as opposed to a rapid response and diminishing decay profile. And instead of the dimensional transformation behavior in phasics, tonics develop a linear sign inversion or opposing signal in response to their stimulus input.
Like their phasic counterparts, tonic aggregates do not convey information about the duration of a stimulus. Tonic aggregates developed from a need to track and characterize changes in their stimulus input. Again, the discussion must emphasize that their behavior is not necessarily a product of their connectivity, and in all probability, the very first tonic circuits were just a simple feedback of a sign inversion.
From a design point of view, tonic assemblies can be simplified as “resonate” neural coalitions. By this we say that in a very simplified description, the activation of a certain tonic coalition (by whatever type of preceding neural activity), the “stimulus” activates another neural ensemble, whose response is subsequently connected back to and reactivates the original coalition, which again activates the second ensemble in a cyclic, mutually activating process.
Since the activation of the second ensemble must by definition be an input to the first coalition in order to reactivate it, this input can act as a form of feedback, if it is tuned to, or “resonates” with the preceding neural “stimulus” that activated the tonic assembly at the very beginning of the cycle. Note that this feedback is the only component characterized by connectivity. The “stimulus” to the first coalition sets the tone of the feedback aggregate and the activation of the second ensemble maintains it. This feedback not only maintains the cycle reaction of the total aggregate, but can also modulate the “tone” of the cycle, by implementing a generated sign inversion in the modulation of its response to changes in the input stimuli, the so-called “negative feedback cycle”. This modulation can thereby establish a desired, balanced state in response to linear changes in the original stimuli.
Indeed, as has been pointed out by many, the tonic assembly form which led to homeostatic processes were themselves a product of the very primal technique employed by the genome in its time-honored process of acquiring and storing genetic successes. Additionally, Nature added the need for the aforementioned built-in response delay in feedback assemblies, to compensate for any mismatch in the response dynamics of the feedback compared to changes in the input stimuli, which results in overshoot and other dynamic imbalances.
Feedback loops are employed extensively throughout most every structure in the human brain, and they are a useful mechanism to accomplish many neural processing tasks. They are so versatile because the basic feedback circuit can be implemented with a very small number of neurons, and it allows much larger assemblies that exhibit more complex behavior.
And it is not hard to envision Nature building even more sophisticated circuits with the modular combinations of phasic and tonic assemblies possible, perhaps with phasic assemblies feeding tonic feedback loops, or multiple feedback loops performing spatial and temporal control functions. And these assemblies would fit in very nicely into a hierarchical engineers’ toolbox. But the natural environment holds many surprises for the hierarchical designer, as Nature must have discovered even at the very beginning of her experimentation with these basic, primal neural assemblies. And although somewhat speculative, these early lessons that Nature discovered were undeniably the most significant, for they would set the mold for the evolution of all neural assemblies to come.
In order to gain a better perspective on this experimentation, we would want to ask how even the most primitive of neural complexes came about. So the dialog will ask the reader to accompany the discussion as we go back in time to see the many primitive heterotrophic organisms that Nature was evolving, in which forms of motility had just begun the division of labor between nerve cells and muscle cells to fashion neural arrays directly connecting the metabolic processes of these metazoans to their developing motility, and increasing the organisms chances of gaining energy.
If we were to observe the environment surrounding any of these newly motile metazoans, certainly we would not find it to be just a single gradient of organic compounds, merely available for the organism to simply motor through. For sure, any environment surrounding our primordial metazoan, whether it be aqueous or terran (indeed, the conjectured environment of pre-Cambrian and Cambrian life was a combination of both, at the bottom of shallow coastal waters), would conceivably present innumerable obstacles to blind locomotion and multiple degrees of various organic gradients. Since Nature rarely produces an organism with locomotion that does not have some mechanism for sensing those organic gradients that are necessary for its survival, this illuminates one of the earliest selection pressures in primitive metazoan nervous array evolution – the integration of environmental sensation with organic motility to even further multiply an organisms chances of gaining energy.
This selection pressure served to evolve many primordial organisms along two parallel lines – a structural evolution which shaped and streamlined the organic body into a singular axis to maximize locomotion in that direction, and the evolution of a sensory structure that would transduce physical events in the environment into neurological signals to better allow the organism to steer the locomotive axis in a direction based on the immediate presence of environmental obstacles and gradients.
These sensory structures may have begun as simple receptor cells embedded in the exolayer of the organism, cells possessing chemical processes to which certain free floating molecules in the environment would bind to, creating a specific electro-chemical reaction. These receptor cells, tuned for an affinity to those molecules which accompany the presence of certain organic compounds, also possessed neural appendages which transmitted a neural impulse triggered by this electro-chemical reaction to the organisms’ developing nervous array. Additional sensory structures would have developed to “sense” the motion of the surrounding medium, and more importantly, structures to signal the immobility of surrounding obstacles, and perhaps primitive transducers for heat and vibration. (The dialog uses the term “sense” here very loosely, since at this level of complexity, there is no abstraction occurring in these neural structures, only a one-to-one correspondence between the triggering of an electro-chemical event and the simple signaling of that event to the neural array.)
With these simple structures, the evolving neural arrays in primitive organisms would have new mechanisms to begin organizing the developing phasic and tonic assemblies that Nature was beginning to experiment with into integrated aggregates that might provide true information of the environment to the organism, gaining abilities for its neural array to better direct the steering function of its co-evolving locomotive axis.
The selective pressure to integrate the effective motoring neurons with the information gathering affective sensory neurons served to develop the most fundamental level of organization for the evolving neural array (and its subsequently more complex versions of central nervous systems), a level of organization that the Organon Sutra has termed the orientation stage in massively asynchronous assemblies and neural systems. This stage will set the foundation for all neurological development to follow in the subsequent evolution of self-directed organisms.
Now, for purposes of discussion, let us pursue an imaginative scenario in order to envision the progression as this primitive sensory/motoric integration of the neural array might have evolved in our primordial metazoan, without any claim that natural evolution actually followed this path, although it is not too much of a stretch to think that Nature would have truly developed extant organisms in this fashion.
Additionally, as bottom-up engineers, we want to view this early evolution from the perspective of its developing neural array, with a focus on those principles which will form our working design doctrine for the much more complex central nervous systems to come, a design doctrine that the Organon Sutra has so far referred to as the formalisms of massively asynchronous assemblies, a doctrine which will guide us in our engineering of the emergent behaviors we have been seeking. As bottom-up engineers, it is our desire to learn those same lessons in nervous system evolution that Nature herself discovered, and as we shall shortly discover, we must trace back to even this primordial metazoan evolution to learn what Nature first brought into existence only after generations and generations of experimentation.
We have begun using the term neural array to refer to the primitive neural assemblies which have yet to evolve into a complexity of even the reptilian brain introduced in discussion 9A, although they have developed simple neural effectors which, by some as-yet defined stimulus, can stimulate a basic motoric system capable of movement along its singular locomotive axis. And for purposes of discussion, we will establish a basic segregation of both the anatomy and the functionality of the evolving neural array, even though our thought exercise has yet to imagine any components beyond the neural motoric effectors. This segregation will divide all evolving neural arrays along the classic three sections, with a section comprising a sensory input complex which translates the impulses arriving from the electro-chemical processes on the exolayer of the organism into signals representative of organic gradients and environmental obstacles, and an intermediate complex to match the metabolic states of the organism with those representative signals of the environment to produce impelling signals for motility, and finally a motoric complex to execute those impelling signals.
With the definition of these segregations, it would be tempting to adapt a hierarchical mindset, surely permissible now at such a primitive stage of neural development, and assert a “direction of flow” for the signaling throughput within the neural array, starting with the translation of environmental signals, flowing down to their reconciliation with the neural activity signaling the organisms metabolic state, followed by the “obvious” determination of steering signals for the locomotive axis.
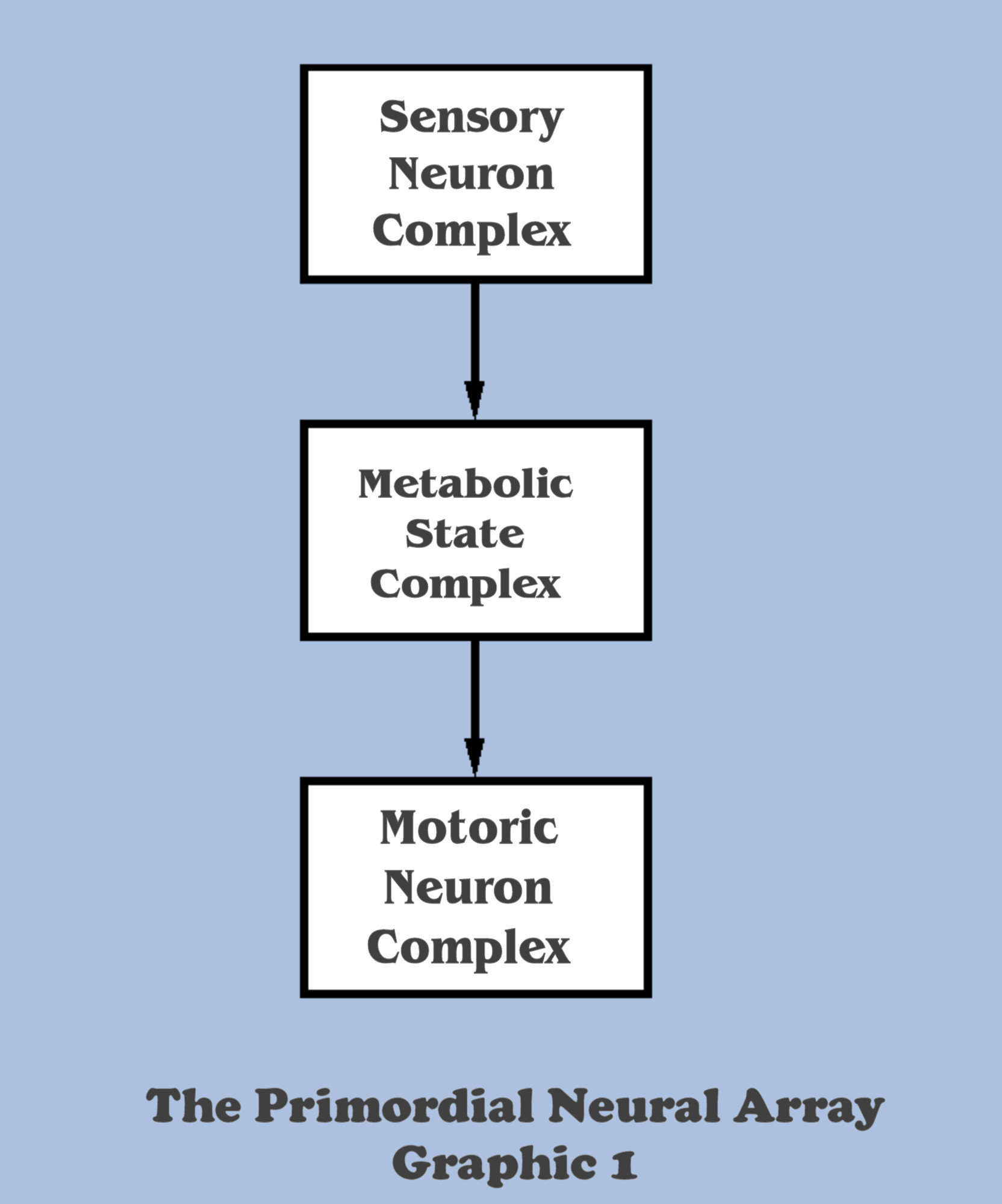
However, we will shortly discover the same lesson that Nature must have learned those many millennia ago, the lesson that neural functionality must begin with a consideration of motoric imperatives first, and only then can the design consider the metabolic requirements which might be satisfied by the existentials of the immediate environment as signaled by the sensory complex. Nature discovered “bottom-up” engineering from the very beginning.
And yet, this thinking runs entirely counter to the intuitive organization of Graphic 1, so surely the hierarchically minded would point out the backward nature of that design perspective and stress the difficulty of developing motoric imperatives before the accumulation of information regarding environmental conditions. How can a neural array develop commands for the locomotive axis before discerning the sensory environment that will form the command parameters in the first place? What is this developmental phenomenon that has the hierarchically programmed design approach turned upside-down?
If we continue our imagination scenario of the development in primal neural array evolution, but focus on the functionality that the motoric complex would likely develop, we can perhaps observe that there are certain generalities that Nature would be likely to embrace.
As the structural evolution was streamlining the body of our organism (species) into a functional locomotive axis, the neural directives necessary to direct this locomotive axis might seem to generalize into three basic, mutually exclusive commands: Go Forward, Go “Left”, and Go “Right”. Now, for sure, the designations of “Left” and “Right” are relative, and perhaps but not necessarily orthonormal to the organisms dorsal vector (and we could easily substitute the nautical terms ‘port’ and ‘starboard’!), and certainly for organisms evolving in a three dimensional aqueous environment, there are “up” and “down” components to any command steering the organisms’ locomotive axis. But as we proceed with the imagination scenario, the interested reader will undoubtedly conclude that these details have little influence on that first important lesson that Nature herself had to learn, a lesson for which she is still seeking solutions to, in even present day extant species, and a lesson which should focus the entire mindset of the bottom-up engineer by turning hierarchical thinking upside-down.
Although they seem simple enough, there is a fundamental dilemma with these three, generalized steering commands that follow from a hierarchical perspective, and not because they are overly generalized. As our evolving organism (species) motors through the environment, steering its locomotive axis in our imagination scenario, it will encounter what Nature must have also generalized as two polarized environmental gradients.
Within our imagination exercise, we can label those two distinct, environmental gradients as aversive conditions and attractive conditions. Aversive conditions are those environmental gradients that our organism will want to steer away from, and attractive conditions are those environmental gradients that our organism will want to steer toward.
Now, it should be apparent to even the most cursory reader that the motoric imperatives of “steer away” and “steer toward” are on a very different order than the functionally minded “steer left”, “steer forward” and “steer right” commands. There is no neat, logical flow from the aversive/attractive imperatives toward a decisive directional axis command, no straightforward hierarchical connection between emotive polar imperatives and logical directional orders.
As scientists and engineers, we can only wonder how Nature first came to reconcile these two dichotomous command levels. And yet, for all motile organisms, survival demanded that reconciliation.
Natures’ solution to that reconciliation could only come about after the recognition that the imperative emotive decision must be made before a logical direction decision can be made. The organism must decide whether an encountered gradient is aversive or not before deciding on a directional strategy to take. And the significance of this priority was emphasized by another salient aspect. The bottom-up engineer should keep in mind that this overriding emotive decision wholly bypasses the neural complex which is signaling the metabolic state of the organism, making it a decision which must go directly from the sensory complex to the motoric complex. Aversion is aversion no matter how hungry or sated an organism is…
It is this “priority command” aspect which creates the first wrinkle in the top-down functionality of neural arrays. Since the emotive decision of aversion/attraction had to be made before any logical direction decision could be made, survival placed a premium on the ability to make that emotive decision, because the organism that could make this determination the quickest was the most likely to survive. And so it should be no surprise to the bottom-up designer that the selective pressures for this specialization would evolve a fourth neural complex in our developing neural array, the emergence of the emotive complex.
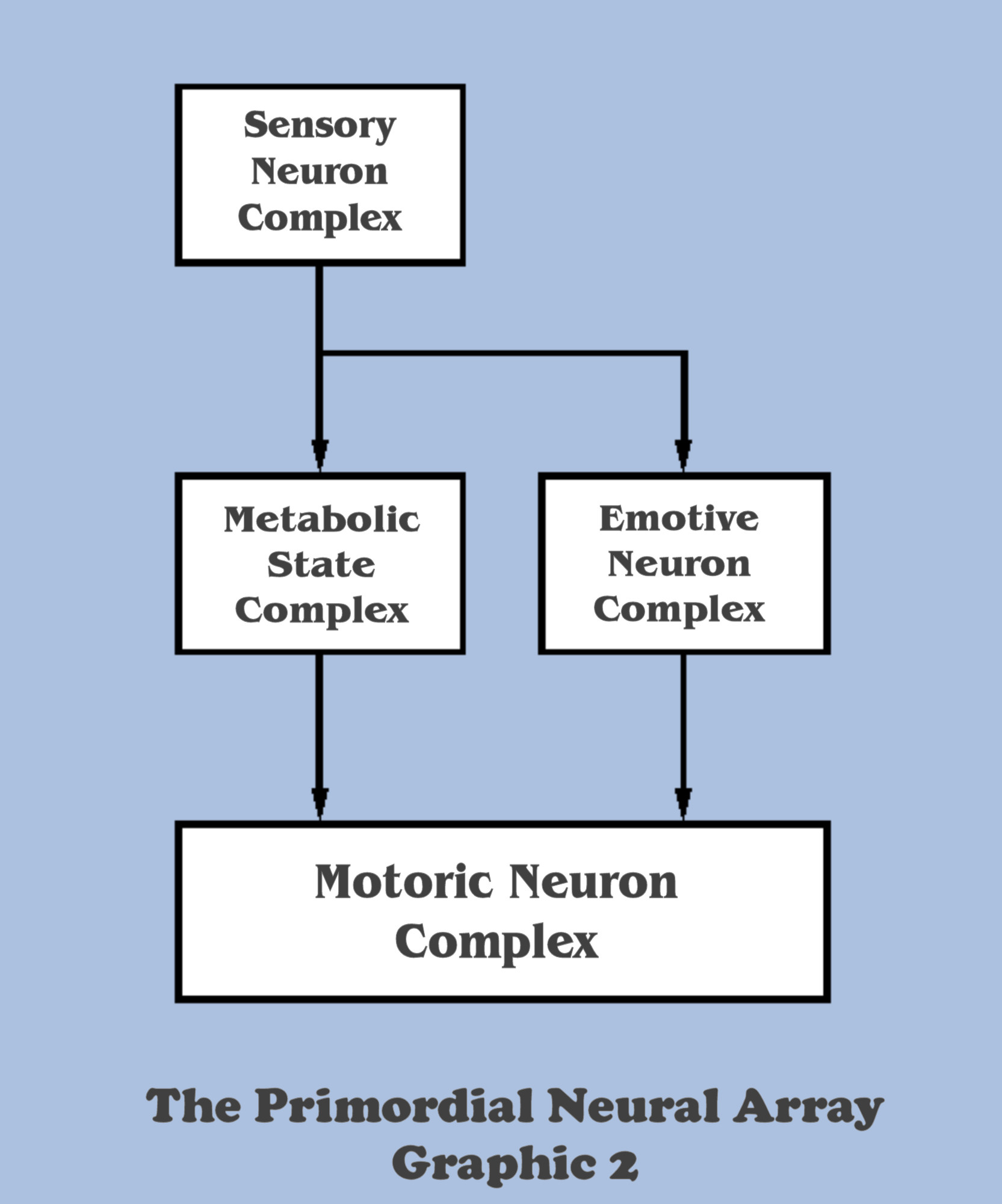
This new complex created, for all time, a schizophrenic aspect to the behavior of all mobile organisms, because the bottom-up engineer should take note that the motoric complex now has two separately derived inputs for the determination of mobility orders. (And with that, after begging a thousand pardons, this dialog cannot help but to throw in the quip that even at this most primitive stage in their evolution, our primordial organisms are already candidates for the psychiatrists’ couch…).
All levity aside, there are profound implications for such a drastic reorganization of even the most basic complexity in neural arrays at such an early stage in their evolution, an adaptation which was demanded by selection pressures even before the specializations that the “top” sensory complex could develop. Many of those implications will be explored in this dialog, as they are all critical to a path to artificial intelligence, but again, significant foundations must be laid first. However, there remain some compelling aspects of this evolutionary development which do need to be taken up at this point, because the development of the emotive complex is only half of the lesson that Nature was learning in her reconciliation of intentional motility.
And hopefully, the bottom-up engineer is gaining a clearer understanding of that lesson at this point, a lesson which can be summarized by acknowledging that motility is no longer a simple matter of seeking a goal that is desired or which has been discovered in the environment, as so many AI researchers fatally simplify it as. Because that environment also presents dangers to an organisms’ survival, the organism must be forever intentional in its locomotion, always balancing aversion with goal attainment as it phylogenetically develops motility schemes. In contrast, organisms that do not exhibit intentional motility are those protozoa and lower metazoan possessing a simple inventory of behavior patterns limited to the singular avoidance/attraction response to environmental gradients. Mobile organisms can no longer afford to be passive occupants in the environment they inhabit, survival demands that they become an active agent in that habitation.
However, before we do approach the next part of the lesson, the dialog will defer to those readers who might be insistent that the discussion cannot abandon so quickly the irrational or incoherent potential posed by a motive system driven by separate command complexes, even though that conversation will require the introduction of several concepts for common discourse. Although we do not want to jump to a different act in the play, the insistent reader can be assured that Nature would find a solution to dual command inputs by fostering the adoption of what ethologists generalize as “releasing mechanisms” – those neurophysiological processes that serve as functional “book ends” to motility strategies that prevent incoherent motility behavior, phylogenetically engineered strategies that structure the balance between emotive and logical motility controls. In present day mammals these “releasing mechanisms” are expressed in the neural structure termed the Basal Ganglia, but again, the story of its evolution is a different act in the play. Studies have confirmed that this structure is an evolutionarily ancient part of the brain, evolving before the limbic and cerebral cortex structures altogether, but those adaptations are part of the plot for the discussion on the Third Fundamental Precept.
But it will be a somewhat convoluted path to the emergence of these mechanisms, because the next part of that path lies with the other half of the lesson that Nature was to learn regarding the initiation of any organic motility beyond the simple behaviors of protozoa and lower metazoa, a part of the lesson that would constitute the second wrinkle in any hierarchical, “top down” functional approach to neural array development.
The dialog has said little about the evolution of the sensory complex in our imagination scenario up to this point because any adaptation that took place before the specialization of the emotive complex would certainly have been eclipsed by the sensory adaptations required after the appearance of that complex.
And like many adaptations guided by selective pressure we have seen so far, these changes, necessitated by intentional motility, occurred along two somewhat interrelated tracks, as both tracks involved the subsequent evolution of the sensory complex.
Just as the selection pressures to specialize the determination of emotive steering signals were evolving the emotive complex, those same influences were adapting the sensory complex to develop those electro-chemical processes which could signal an aversive condition as efficiently as possible. And although there will be much to say about this phylogenetic process in the next discussion, the current discussion will focus on the much more important second track of the sensory complex adaptation.
The exigencies which evolved the emotive complex in those developing neural arrays in turn created another functional necessity which would influence the adaptation of the neural arrays’ sensory complex also. Once survival placed a premium on the mechanism that determines the presence of an aversive gradient in the first place, Nature discovered that organisms which simply motored on in a perpetual avoidance mode were not all that survivable.
As mentioned, when an organism encounters an attractive gradient in the environment, the motility strategy required is not very complex at this level of neural development. The organism simply moves through the gradient, making left or right adjustments in the locomotive axis that maximizes the degree of gradient. And indeed, the only element which logically ties together the complexes of Graphic 1 in a hierarchical manner was the functionality of the metabolic state determining the inception and extinction of any motoric signal toward an attractive gradient.
But after encountering an aversive gradient, this hierarchy becomes broken, as the new mobility strategy becomes more complex. Certainly, the organism does not want to continue moving on as before it encountered the aversion, but “do not go forward” is not a motoric command that has any meaning for a neural complex controlling a locomotive axis that has only three “settings” of forward, left or right. A mitigation strategy needs to “know” more. Go left or go right? And for how far? And just as importantly, when can the organism signal the extinction of the aversive condition and return to any goal attainment strategies. There is a whole chain of events required here, and each event in the chain is dependent on the successful completion of the event preceding it. Whereas steering in an attractive gradient is not so much of a strategy, steering in an aversive environment most certainly is.
The selection pressure to develop mitigation strategies would not only shape the functionality of the developing emotive complex in the neural array of organisms, but the “need to know more” would also direct the adaptation of the sensory complex to develop electro-chemical processes and neural “translators” which might signal distinct environmental cues, triggering mitigation strategies having higher survival value than others. Returning to our imagination scenario, it is not hard to see how, in this evolutionary process, the emotive complex is driving the adaptation of the sensory complex, from the “bottom-up”, and even developing true neural feedback mechanisms which serve to “fine-tune” the sensory complex and allowing the selection of those varying environmental cues as a mitigation strategy is executed.
This will become a most significant and recurring theme as Nature, and those students of Massively Asynchronous Assemblies will come to learn, that the formation of all feedback mechanisms in a neural array occurs from the “bottom-up”.
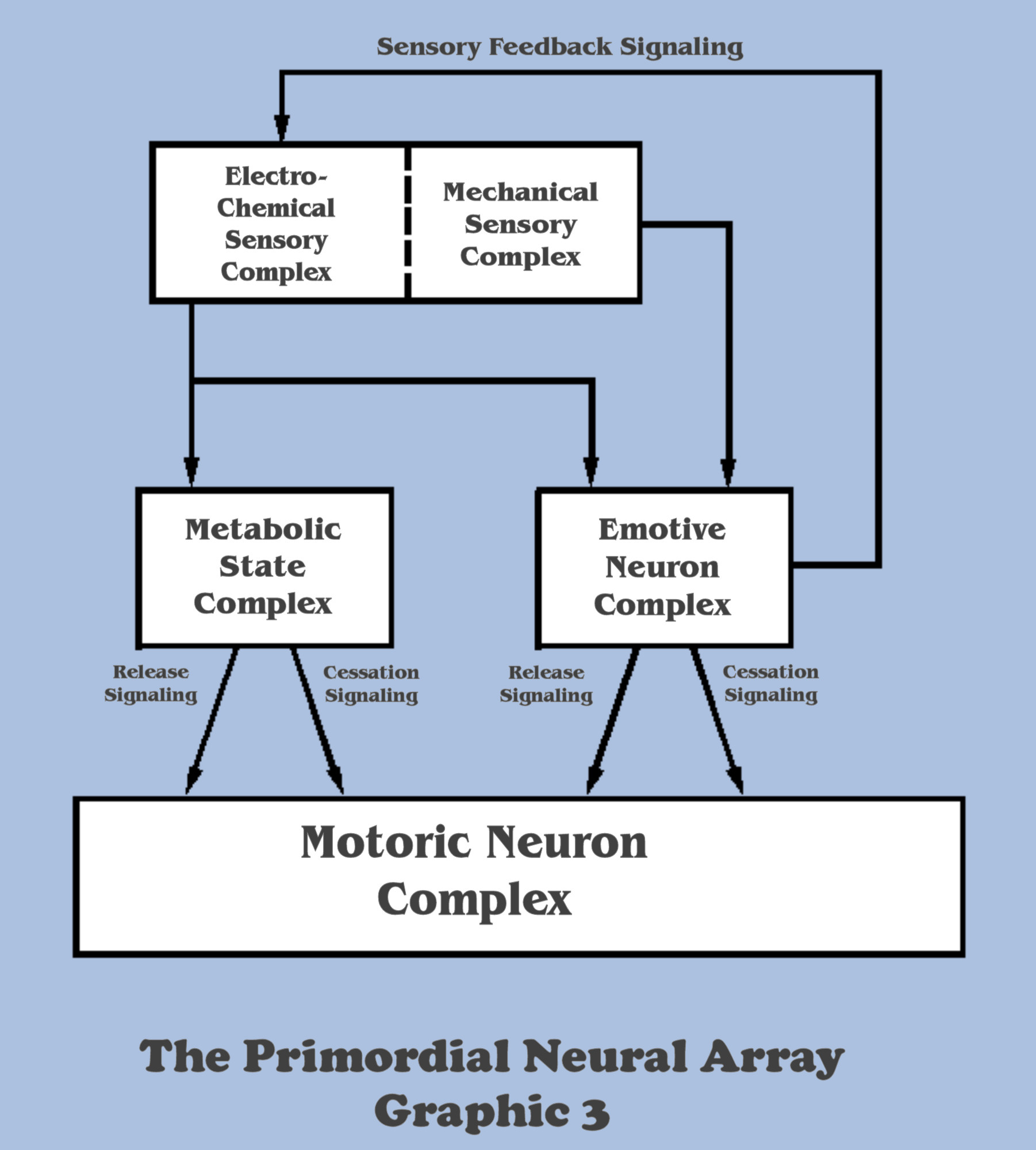
For this reason, there will forever be no hierarchy from sensory affection to motoric effection discernable because of the intimate, interdependent co-evolution between the neural sensory complex, the emotive complex and the continuous feedback between them, no matter how many layers of neural complexity evolution will form around this core of functionality. This is why it is meant that the functionality of the motoric complex in any neural array will be the first impetus for the selective evolution of the neural sensory complex, and why that aspect should forever banish considerations of hierarchical thinking from the mindset of the bottom-up engineer.
And in fact there was one such emotive evolution which probably contributed to the success of the most prolific creature to live in the Cambrian era. The trilobites were among the most successful of all early arthropods, from which the crustaceans, and later the vast number of insect species evolved. Beginning some 520 million years ago, over four thousand species of trilobites diversified throughout many of the Earths’ primordial oceans during a span of 270 million years. And primitive trilobites, having developed their characteristic locomotive axis with a motoric framework based on articulated segments, crawling across the ocean bottom, developed the ability to curl up into a ball, exposing only the armored surfaces of their segmented bodies to threatening predators. This unique motoric sequence, not so much more than a reflex action, placed minimal demands on neural sophistication, but allowed the trilobites to react immediately to threatening situations.

Unfortunately, not all organisms would be availed with such convenient defense mechanisms. Survival in a world teeming with dangers would demand an ever more sophisticated emotive complex as organisms, and their predators, evolved.
So, in addition to recognizing the intentional aspects being developed in the evolving motoric complex, it is just as important for the bottom-up engineer to see that Nature learned even back at this primordial stage that sensory perception is also not just about the sensation of the entire environment, but about the apprehension of intentional aspects of the environment. And those intentional aspects are vectored by the emotive/logical balance in the motoric complex. Because of this, the environment should “look” differently to the sensory complex depending on where the balance is in aversive/attractive control of the motoric complex.
The feedback that is vectoring this diffraction in sensation is essentially a modulator to perception, and this paradigm will follow the evolution of nervous assemblies from this primordial orientation stage of neural arrays all the way to Natures’ glorious expression of modern primate and human central nervous systems.
And this paradigm is one of the central tenets of the Organon Sutra.
And this paradigm requires moving from sensation to elementary abstraction. The selective pressure to develop elementary abstraction behaviors in the perception mechanisms of organisms began even as far back as this primordial orientation stage of neural arrays, and not with the more fully developed, multi-layered complexity of later central nervous systems as is usually characterized. Environmental diffraction, into aversive and attractive conditions, was the first imperative of the sensory complex even before environmental objectification. Survival of the organism simply demanded this priority.
In most mammalian cerebral nervous systems today, we can see this diffraction of perception in the functionality of the thalamus, and it is important for the bottom-up engineer to understand how Nature moved from the basic sensation of aversive and attractive gradients in the primordial neural array to the exquisitely fine-tuned functionality of the thalamus and the particular feedback structures between it and the basal ganglia, and how this sensory diffraction became the common denominator in the equation defining abstraction. Because only 20% of the direct signals to the thalamus come from the sensory structures, with the majority of the remaining inputs formed by reciprocal connections from the cortex, basal ganglia, and other structures, the thalamus should be thought of more than just a sensory relay assembly. Certainly, the dialog will necessarily devote extensive discussions to this critical concept of environmental diffraction.
Additionally, because the stimulus that signals the extinction of an aversive motility strategy originates in the environment, whereas the stimulus that signals the extinction of a goal directed motility strategy originates in the metabolic complexes of the neural array, this paradigm will always be more than an “interpretation” of environmental sensation.
Many researchers see the information -> knowledge reflex as the mechanism to initiate movement, and do not give importance to the mechanisms that cease movement. It is the duality of these inseparable neural duties in which learning also plays a part, but because of that duality, learning must be spread across whole populations of neurons, and no single neuron (or synapse, for that matter) can span that duality in achievement of the learning edict. This is the reason why the bottom-up engineer must be specific before attaching “information” to components at the neural or synaptic level. The mechanistic basis of that learning begins the formation of a feedback loop bridging those complexes that initiate movement with the complexes that extinguish it. But since there is an interdependent chain of processes between initiation and cessation, a concatenation of causalities, the distance between initiation and cessation is not a simple one.
So, first Nature had to develop the ontogenetic mechanisms which would ultimately express this feedback, and although we are even just now (in our primordial imagination scenario) seeing Nature experiment with tonic feedback assemblies within the neural array, recall that this new feedback will have to modulate the sensory complex, a neural assembly that is exposed to the environment. And like a harsh taskmaster, there is no end to the lessons that the environment holds for Natures’ evolution of centralized nervous systems, an environment that is the very embodiment of inexhaustible variety.
Throughout her evolution of centralized nervous systems, the circuits that Nature was developing in the phasic form would in time become the basic building blocks for sensory induction, and the tonic circuits exhibiting resonance would become the basic building blocks for metabolic control, and more importantly, the elementary abstraction of sensory information, but there were more lessons in store for Nature as she tinkered with neural complexity.
The road to a fully developed nervous system was not very straightforward. There are complexities needed beyond the basic functionality of phasic and tonic assemblies. Although it is a matter of speculation, before Nature could concoct even more expressive structures with these two basic forms, a funny thing happened on the way to intelligence.
Nature bumped into nonlinearity.
GO TO NEXT DISCUSSION
GO TO TOP OF DISCUSSION
Copyright © 2019 All rights reserved